แปลและเรียบเรียงจาก: Cinelli, M.A.; Do, H.T.; Miley, G.P.; Silverman, R.B. Inducible Nitric Oxide Synthase: Regulation, Structure, and Inhibition. Med. Res. Rev. 2020, 40(1), 158–189. doi:10.1002/med.21599 (เผยแพร่ผ่าน PMC/HHS Public Access)
คำศัพท์และคำย่อที่ควรทราบก่อนอ่าน
|
คำย่อ |
ความหมาย |
|
iNOS |
Inducible Nitric Oxide Synthase — เอนไซม์สังเคราะห์ไนตริกออกไซด์ชนิดเหนี่ยวนำได้ |
|
nNOS |
Neuronal Nitric Oxide Synthase — เอนไซม์สังเคราะห์ไนตริกออกไซด์ชนิดพบในเซลล์ประสาท |
|
eNOS |
Endothelial Nitric Oxide Synthase — เอนไซม์สังเคราะห์ไนตริกออกไซด์ชนิดพบในเซลล์บุผนังหลอดเลือด |
|
NO |
Nitric Oxide — ไนตริกออกไซด์ |
|
L-Arg |
L-Arginine — กรดอะมิโนแอล-อาร์จินีน (สารตั้งต้นของ NO) |
|
H4B |
(6R)-5,6,7,8-Tetrahydrobiopterin — โคแฟกเตอร์สำคัญของเอนไซม์ NOS |
|
NADPH |
Reduced Nicotinamide Adenine Dinucleotide Phosphate — โคเอนไซม์ให้อิเล็กตรอน |
|
FAD / FMN |
Flavin Adenine Dinucleotide / Flavin Mononucleotide — โคแฟกเตอร์ฟลาวินในสายพาอิเล็กตรอน |
|
CaM |
Calmodulin — โปรตีนจับแคลเซียมที่ควบคุมการทำงานของ NOS |
|
LPS |
Lipopolysaccharide — สารพิษจากผนังเซลล์แบคทีเรียแกรมลบ ใช้เหนี่ยวนำการอักเสบ |
|
cGMP |
Cyclic Guanosine Monophosphate — สารสื่อสัญญาณระดับเซลล์ที่เกิดจากฤทธิ์ของ NO |
|
ONOO⁻ |
Peroxynitrite — เปอร์ออกซีไนไตรต์ สารออกซิแดนท์ที่รุนแรงเกิดจาก NO รวมกับซุปเปอร์ออกไซด์ |
|
IC50 / Ki / Kd / EC50 |
ค่าตัวเลขบ่งชี้ความแรงของสารยับยั้งหรือสารออกฤทธิ์ ยิ่งค่าต่ำยิ่งมีฤทธิ์แรง |
|
i/e, i/n |
อัตราส่วนความจำเพาะของสารยับยั้งต่อ iNOS เทียบกับ eNOS (i/e) หรือ nNOS (i/n) ยิ่งค่าสูงยิ่งจำเพาะต่อ iNOS มากขึ้น |
|
SAR |
Structure-Activity Relationship — ความสัมพันธ์ระหว่างโครงสร้างเคมีกับฤทธิ์ทางชีวภาพ |
เมื่อร่างกายสร้าง “อาวุธก๊าซ” ขึ้นมาต่อสู้กับเชื้อโรค
ลองนึกภาพสงครามที่เกิดขึ้นทุกวินาทีภายในร่างกายของเรา เมื่อแบคทีเรียบุกรุกเข้าสู่กระแสเลือด เซลล์เม็ดเลือดขาวชนิดแมคโครฟาจจะเปลี่ยนตัวเองให้กลายเป็นโรงงานผลิตอาวุธเคมีขนาดจิ๋วอย่างรวดเร็ว อาวุธที่ว่านี้คือก๊าซพิษชนิดหนึ่งที่ชื่อว่า “ไนตริกออกไซด์” (nitric oxide หรือ NO) ซึ่งสามารถทำลายเชื้อโรคได้อย่างมีประสิทธิภาพ แต่หากผลิตออกมามากเกินไปหรือควบคุมไม่ได้ ก๊าซชนิดเดียวกันนี้ก็สามารถทำร้ายเนื้อเยื่อของร่างกายเราเองได้เช่นกัน นี่คือเรื่องราวของเอนไซม์ที่อยู่เบื้องหลังกระบวนการนี้ นั่นคือ Inducible Nitric Oxide Synthase หรือ iNOS
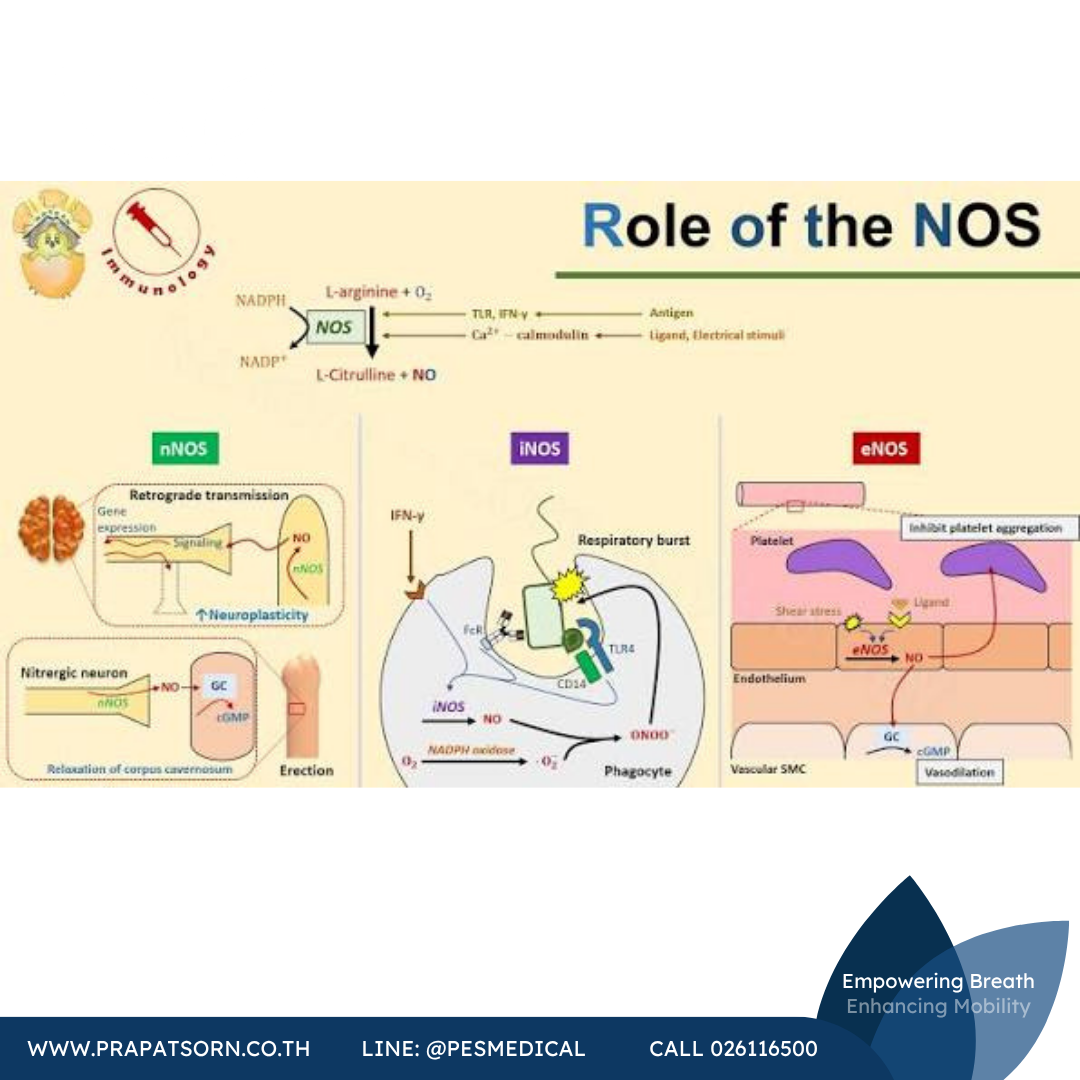
ไนตริกออกไซด์เป็นโมเลกุลส่งสัญญาณระดับเซลล์ที่สำคัญ มีบทบาทในกระบวนการทางสรีรวิทยาที่หลากหลายของสัตว์เลี้ยงลูกด้วยนม ทั้งการขยายหลอดเลือด (vasodilation) การคลายตัวของกล้ามเนื้อเรียบ การส่งสัญญาณประสาท และการตอบสนองของระบบภูมิคุ้มกัน NO เป็นอนุมูลอิสระ (free radical) ที่ถูกสร้างขึ้นโดยกลุ่มเอนไซม์ที่เรียกว่าไนตริกออกไซด์ซินเทส (nitric oxide synthases หรือ NOSs) ผ่านการออกซิไดซ์กรดอะมิโนแอล-อาร์จินีน (L-arginine) ให้กลายเป็นแอล-ซิทรูลลีน (L-citrulline) เอนไซม์ NOS มีอยู่สามไอโซฟอร์มหลัก สองชนิดแรกคือ neuronal NOS (nNOS) และ endothelial NOS (eNOS) เป็นเอนไซม์ที่ถูกแสดงออกอยู่ตลอดเวลา (constitutively expressed) ส่วนชนิดที่สามคือเอนไซม์ที่ถูกเหนี่ยวนำให้แสดงออก จึงเรียกว่า iNOS
nNOS พบเป็นหลักในระบบประสาทและจำเป็นต่อการส่งสัญญาณของเซลล์ประสาท ส่วน eNOS อยู่ที่ชั้นเยื่อบุผนังหลอดเลือดและมีความสำคัญต่อการขยายหลอดเลือดและการควบคุมความดันโลหิต ไอโซฟอร์มทั้งสองนี้ผลิต NO ในปริมาณระดับนาโนโมลาร์เป็นช่วงเวลาสั้นๆ (เพียงไม่กี่วินาทีถึงไม่กี่นาที) โดยอาศัยกลไกที่ขึ้นกับแคลเซียม/แคลโมดูลิน (calcium/calmodulin) ในทางตรงกันข้าม iNOS จะไม่ปรากฏอยู่ในเซลล์ตลอดเวลา แต่จะถูกแสดงออกก็ต่อเมื่อเซลล์ถูกกระตุ้นหรือเหนี่ยวนำ โดยทั่วไปมักถูกกระตุ้นด้วยไซโตไคน์ที่ก่อการอักเสบ (pro-inflammatory cytokines) และ/หรือไลโปโพลีแซคคาไรด์จากแบคทีเรีย (bacterial lipopolysaccharide หรือ LPS) เมื่อถูกเหนี่ยวนำแล้ว iNOS จะผลิต NO ในปริมาณมาก (ระดับไมโครโมลาร์) และคงอยู่จนกว่าเอนไซม์จะถูกย่อยสลาย ซึ่งอาจนานหลายชั่วโมง
ปริมาณ NO จำนวนมากที่ถูกผลิตขึ้นนี้ช่วยป้องกันร่างกายจากเชื้อโรคที่บุกรุกเข้ามา จึงมีความสำคัญอย่างยิ่งต่อการตอบสนองต่อการอักเสบและระบบภูมิคุ้มกันโดยกำเนิด (innate immune system) แต่ในทางกลับกัน หากความเข้มข้นของ NO สูงเกินควรอันเนื่องมาจากการแสดงออกมากเกินไปหรือการควบคุมที่ผิดปกติของ iNOS ก็อาจก่อให้เกิดผลพิษต่อร่างกายและมีความสัมพันธ์กับโรคของมนุษย์หลายชนิด ทั้งภาวะช็อกจากการติดเชื้อในกระแสเลือด (septic shock) ความผิดปกติของหัวใจ ความเจ็บปวด เบาหวาน และมะเร็ง ฤทธิ์สองด้าน (ทั้งเป็นประโยชน์และเป็นโทษ) ของ NO ที่เกี่ยวข้องกับ iNOS นี้ขึ้นอยู่กับความเข้มข้นเป็นสำคัญ ดังนั้นการควบคุมการผลิต NO จึงมีความสำคัญทั้งต่อการรักษาหน้าที่ทางสรีรวิทยาปกติและการควบคุมผลเสียที่อาจเกิดขึ้น
ด้วยความสำคัญเช่นนี้ งานวิจัยเกี่ยวกับ iNOS จึงเป็นสาขาที่ได้รับความสนใจอย่างมาก การสืบค้นคำว่า “iNOS” ในฐานข้อมูล SciFinder พบเอกสารอ้างอิงมากกว่า 37,000 รายการ บทความทบทวนวรรณกรรมฉบับนี้นำเสนอภาพรวมทั่วไปของ iNOS โดยเน้นที่โครงสร้าง หน้าที่ และการควบคุมการทำงาน ทั้งในแง่สรีรวิทยาปกติและพยาธิสรีรวิทยา รวมถึงจะกล่าวถึงการยับยั้ง iNOS ในฐานะกลยุทธ์การรักษาที่มีศักยภาพ และความคืบหน้าของการยับยั้ง iNOS ในการทดลองทางคลินิก
โครงสร้างและหน้าที่ของ iNOS
iNOS เป็นโปรตีนของสัตว์เลี้ยงลูกด้วยนมที่มีมวลโมเลกุล 131 กิโลดาลตัน ประกอบด้วยกรดอะมิโน 1,153 ตัว ซึ่งประกอบกันเป็นสองโดเมนหลัก ได้แก่ โดเมนรีดักเทส (reductase) ที่ปลายด้าน C ซึ่งมีโดเมนย่อยจับฟลาวินโมโนนิวคลีโอไทด์ (FMN) และโดเมนออกซิจีเนส (oxygenase) ที่ปลายด้าน N iNOS มีโครงสร้างควอเทอร์นารีแบบไดเมอร์ที่เชื่อมด้วยสังกะสี (zinc-bridged homodimer) ซึ่งทำให้เอนไซม์สามารถเปลี่ยนแอล-อาร์จินีนให้เป็นแอล-ซิทรูลลีนพร้อมกับผลิต NO ไปด้วย กระบวนการนี้ถูกควบคุมโดยสายพาอิเล็กตรอน (electron transport chain) ที่ซับซ้อน ซึ่งอาศัยโคแฟกเตอร์หลายชนิด ได้แก่ NADPH, FAD, FMN, ฮีม (heme) และ (6R)-5,6,7,8-เตตระไฮโดรไบออปเทอริน (H4B) โดยกระบวนการนี้ถูกควบคุมผ่านแคลโมดูลิน (CaM) ซึ่งจับอยู่ในบริเวณข้อพับ (hinge region) ระหว่างโดเมนออกซิจีเนสและโดเมนรีดักเทส
แม้ iNOS, nNOS และ eNOS จะมีความคล้ายคลึงกันของยีน โคแฟกเตอร์ และหน้าที่โดยรวมในระดับสูง แต่ความแตกต่างของโครงสร้างโดเมนก็ทำให้แต่ละไอโซฟอร์มมีความจำเพาะในหน้าที่และการควบคุมที่ต่างกัน iNOS เป็น NOS ของสัตว์เลี้ยงลูกด้วยนมที่มีโครงสร้างโดเมนเรียบง่ายที่สุด ประกอบด้วยเพียงโดเมนรีดักเทส โดเมนออกซิจีเนส และโดเมนจับ CaM เท่านั้น โดยไม่มีห่วงยับยั้งตัวเอง (auto-inhibitory loop) เหมือนที่พบใน eNOS และไม่มีโดเมน PDZ เหมือนที่พบใน nNOS
เอนไซม์ NOS ทุกไอโซฟอร์มมีอะตอมสังกะสีที่ไม่มีฤทธิ์เร่งปฏิกิริยาอยู่ที่บริเวณรอยต่อของไดเมอร์ โดยสังกะสีหนึ่งอะตอมต่อคู่ไดเมอร์จะสร้างกลุ่มซิงก์เททระไทโอเลต (zinc tetrathiolate cluster) จากการที่แต่ละมอนอเมอร์บริจาคซีสเทอีนสองตำแหน่งในรูปแบบ -CXXXXC- การศึกษาทางชีวเคมีพบว่าบทบาทของไอออนสังกะสีคือการส่งเสริมการจับตัวของ H4B แม้ CaM จะเกี่ยวข้องกับ NOS ทุกไอโซฟอร์ม แต่การจับตัวที่แน่นระหว่าง CaM กับ iNOS (อันเป็นผลจากความแตกต่างของลำดับกรดอะมิโนในบริเวณข้อพับ) ทำให้ iNOS สามารถถูกกระตุ้นได้ที่ความเข้มข้นแคลเซียมทางสรีรวิทยาที่ต่ำกว่ามาก (40 นาโนโมลาร์ใน iNOS เทียบกับ 400 นาโนโมลาร์ใน nNOS และ eNOS) เนื่องจากความเข้มข้นแคลเซียมในไซโตพลาสซึมโดยทั่วไปอยู่ที่ราว 100 นาโนโมลาร์ จึงอาจกล่าวได้ว่า iNOS แทบจะอยู่ในสถานะทำงานตลอดเวลาโดยที่การควบคุมผ่านแคลเซียมแทบไม่มีความหมายอีกต่อไป การดัดแปลงหลังการแปลรหัส (post-translational modifications) ก็อาจมีความสำคัญเชิงหน้าที่หรือการควบคุมเช่นกัน ตัวอย่างเช่น มีการตรวจพบฟอสโฟไทโรซีนบนโมเลกุล iNOS และการทดสอบแสดงให้เห็นว่าการยับยั้งฟอสฟาเทสในเซลล์ด้วยวานาเดตนำไปสู่การเพิ่มขึ้นของกิจกรรม iNOS
กลไกการสร้างไนตริกออกไซด์
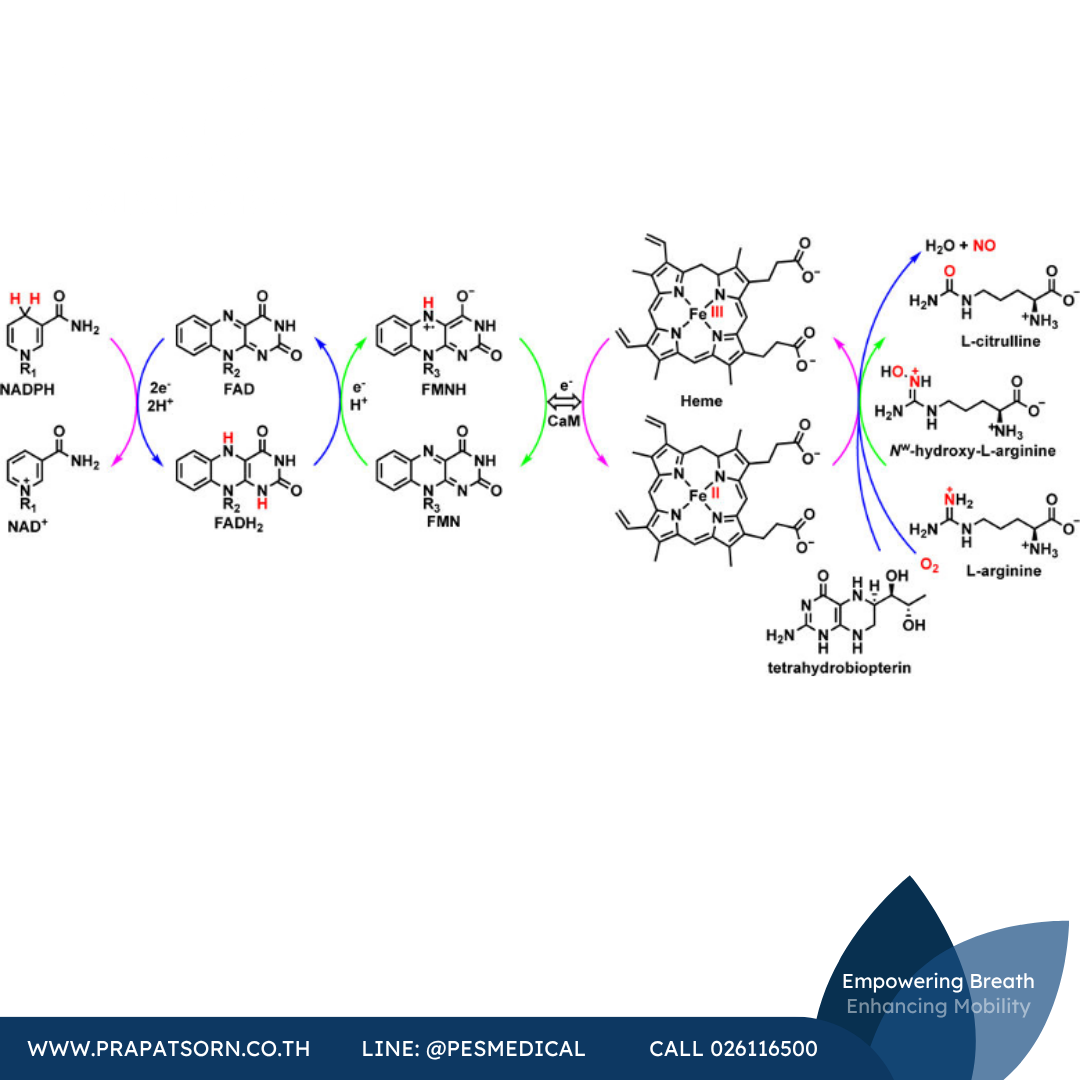
แม้แรงขับเคลื่อนหลักของการสร้าง NO จะมาจาก NADPH แต่วงจรเร่งปฏิกิริยาดำเนินไปผ่านสายพาอิเล็กตรอนภายในและระหว่างโปรตีนที่ซับซ้อน วงจรเริ่มต้นเมื่อ NADPH จับกับโดเมนรีดักเทสและกระตุ้นให้เกิดปฏิกิริยารีดักชันของ FAD ที่อยู่ใกล้เคียงในรูปแบบสองอิเล็กตรอน ภายในโดเมนรีดักเทส FAD อยู่ใกล้กับโดเมนย่อย FMN ซึ่งทำให้เกิดปฏิกิริยารีดักชันหนึ่งอิเล็กตรอนของ FMN โดย FAD ในขั้นตอนนี้ การถ่ายเทอิเล็กตรอนเชิงพื้นที่จากโดเมนหนึ่งไปยังอีกโดเมนหนึ่งยังไม่เป็นที่แน่ชัดนัก เนื่องจากโดเมนย่อย FMN อยู่ห่างจากโดเมนออกซิจีเนสที่มีฮีมอยู่ การศึกษาต่างๆ แสดงให้เห็นความยืดหยุ่นอย่างมากภายในโปรตีนและได้อธิบายการเปลี่ยนแปลงเชิงโครงสร้างระหว่างวงจรเร่งปฏิกิริยา เมื่อ CaM และแคลเซียมกระตุ้นให้เกิดความไว เอนไซม์จะบิดโดเมนย่อย FMN ให้เข้าใกล้กับโดเมนออกซิจีเนสที่มีฮีม ที่น่าสนใจคือโดเมน FMN จะไม่ถ่ายเทอิเล็กตรอนให้ฮีมในมอนอเมอร์ของตัวเอง แต่จะถ่ายเทให้กับมอนอเมอร์อีกตัวหนึ่งในไดเมอร์ ซึ่งอธิบายได้ว่าเหตุใด iNOS ในรูปมอนอเมอร์ (และ NOS โดยทั่วไป) จึงไม่มีฤทธิ์เร่งปฏิกิริยาอิเล็กตรอนที่ได้รับจาก FMN จะทำให้เหล็กในฮีมถูกรีดิวซ์จากเหล็ก(III) เป็นเหล็ก(II) และเตรียมความพร้อมของตำแหน่งเร่งปฏิกิริยาให้สามารถรับและกระตุ้นออกซิเจนโมเลกุลได้
กลไกที่แน่ชัดของการกระตุ้นนี้ยังเป็นที่ถกเถียงกันอยู่ อย่างไรก็ตาม ใกล้กับตำแหน่งเร่งปฏิกิริยามีบริเวณจับ H4B ซึ่งเชื่อว่ามีบทบาทหลายประการในการผลิต NO H4B ทำให้เกิดโครงแบบสปินสูง (high spin) ของเหล็กในฮีม ช่วยรักษาเสถียรภาพของรูปแบบไดเมอร์ของเอนไซม์ และอาจส่งเสริมการเชื่อมโยงของ NADPH เข้ากับการผลิต NO อย่างมีประสิทธิภาพ นอกจากนี้ ในระหว่างการออกซิไดซ์แอล-อาร์จินีน H4B ยังมีบทบาทสำคัญในฐานะตัวส่งและแหล่งกักเก็บอิเล็กตรอนภายในตำแหน่งเร่งปฏิกิริยาอีกด้วย
เมื่อแอล-อาร์จินีนจับกับกรดกลูตามิกที่ตำแหน่งเร่งปฏิกิริยา ฮีมโคแฟกเตอร์จะดำเนินกลไกคล้ายเอนไซม์ P450 เพื่อออกซิไดซ์แอล-อาร์จินีนให้เป็น Nω-ไฮดรอกซี-แอล-อาร์จินีน (L-NOHA) จากนั้นจะเกิดขั้นตอนที่สองซึ่งออกซิไดซ์ L-NOHA ต่อไปเป็นแอล-ซิทรูลลีนและ NO ในสองจุดของกระบวนการเปลี่ยนแอล-อาร์จินีนเป็น NO นั้น H4B ทำหน้าที่ให้ความเสถียรทางประจุระหว่างการกระตุ้นออกซิเจนโมเลกุล การบริจาคอิเล็กตรอนจาก H4B ไปยังสารมัธยันตร์เหล็ก(IV)ไฮโดรเปอร์ออกซิลของฮีม ทำให้เกิดไอออนบวกแบบเรดิคัลออกซีนอยด์เหล็ก(IV) ซึ่งเชื่อว่าเป็นสารมีฤทธิ์เร่งปฏิกิริยา ในขั้นตอนสุดท้ายที่ปลดปล่อย NO ออกจากฮีม H4B จะรับอิเล็กตรอนกลับคืนจากเหล็กหนึ่งตัว ทำให้เกิดขั้วบนพันธะเหล็ก-ไนโตรเจน และเปิดทางให้ NO แพร่กระจายออกจากตำแหน่งเร่งปฏิกิริยาได้ ในระหว่างการเร่งปฏิกิริยา H4B บริจาคอิเล็กตรอนสองตัวแต่รับกลับคืนเพียงหนึ่งตัว แหล่งที่มาของอิเล็กตรอนอีกตัวหนึ่งคือโดเมนย่อย FMN แต่กลไกที่แน่ชัดและการจัดเรียงเชิงพื้นที่ที่จำเป็นในการส่งผ่านอิเล็กตรอนนี้ยังไม่เป็นที่ทราบแน่ชัด
ภาพที่ 1: สายพาอิเล็กตรอนจาก NADPH ผ่าน FAD และ FMN ไปยังฮีม โดยอาศัย H4B ในการเปลี่ยนแอล-อาร์จินีนเป็นแอล-ซิทรูลลีนและไนตริกออกไซด์
การจัดเรียงตัวเชิงพื้นที่ของ iNOS
หลักฐานทางชีวเคมีจำนวนมากในอดีตชี้ให้เห็นว่าอิเล็กตรอนถูกส่งผ่านจากโดเมนย่อย FMN ไปยังฮีมในโดเมนออกซิจีเนสของมอนอเมอร์อีกตัวหนึ่ง แต่จนกระทั่งเมื่อไม่นานมานี้ ก็ยังไม่มีคำตอบที่ชัดเจนสำหรับคำถามพื้นฐานเกี่ยวกับการจัดเรียงเชิงพื้นที่ของโดเมนต่างๆ และการเคลื่อนไหวของโดเมนเหล่านั้นเพื่อให้เกิดการเร่งปฏิกิริยา ปัญหานี้ส่วนใหญ่เกิดจากความยากในการตกผลึกเอนไซม์ NOS แบบไดเมอร์ที่มีความยาวเต็มโมเลกุล อันที่จริง โครงสร้างผลึกของโดเมนออกซิจีเนสแบบไดเมอร์ของทั้งสามไอโซฟอร์มเพิ่งได้รับการไขในช่วงปลายทศวรรษ 1990 และสำเร็จสมบูรณ์ในปี ค.ศ. 2002 ด้วยการตีพิมพ์โครงสร้างของ nNOS ต่างจาก nNOS หรือ eNOS ตรงที่ปฏิสัมพันธ์อันแน่นหนาระหว่าง CaM กับ iNOS ทำให้สามารถตกผลึกร่วมกันได้ (co-crystallization) ซึ่งช่วยยืนยันตำแหน่งของ CaM ในเอนไซม์ NOS ได้ แต่ก็ยังไม่สามารถอธิบายการจัดเรียงโดยรวมได้ชัดเจนนัก
ในปี ค.ศ. 2014 กลุ่มวิจัยของ Marletta ได้ตีพิมพ์การศึกษาด้วยเทคนิค cryo-EM ของเอนไซม์กลุ่ม NOS ซึ่งแสดงโครงสร้างสามมิติของโดเมนต่างๆ ในตระกูล NOS และแสดงภาพรวมของวงจรเร่งปฏิกิริยาแบบเป็นขั้นตอน โดยพบว่า iNOS พับตัวเองเกือบครึ่งหนึ่งเพื่อส่งอิเล็กตรอนจาก FMN ไปยังฮีม แม้ความละเอียดของภาพเหล่านี้จะจำกัดอยู่เพียงระดับโครงสร้างของโดเมน แต่การจำลองทางคอมพิวเตอร์ของรูปทรงสามมิติที่สังเกตได้ก็ชี้ให้เห็นถึงความยืดหยุ่นในระดับสูงของบริเวณจับ CaM รวมถึงความสามารถของโดเมนย่อย FMN ในการเคลื่อนที่เป็นระยะทางไกลได้อย่างอิสระจากโดเมนรีดักเทสที่มันสังกัดอยู่โดยทั่วไป
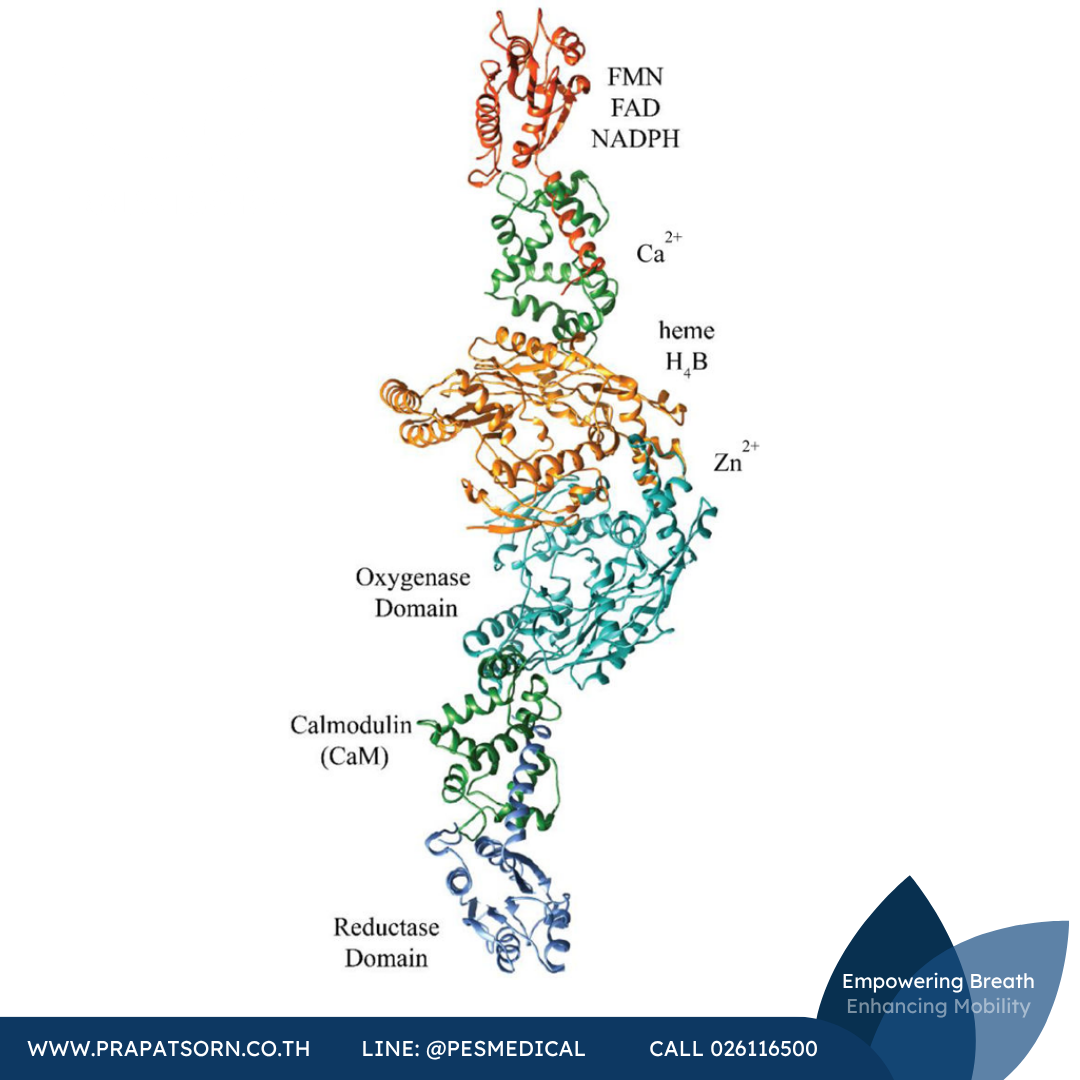
ภาพที่ 2: โครงสร้างผลึกแบบไดเมอร์ของ iNOS ที่ประกอบขึ้นจากข้อมูล cryo-EM แสดงโดเมนรีดักเทส โดเมนออกซิจีเนส แคลโมดูลิน และตำแหน่งของโคแฟกเตอร์ต่างๆ
กรดอะมิโนสำคัญใน iNOS
การวิเคราะห์การกลายพันธุ์ของ iNOS ได้ระบุกลุ่มกรดอะมิโนที่มีความสำคัญต่อการทำงานของเอนไซม์ไว้หลายตำแหน่ง เมื่อ Glu546 ถูกกลายพันธุ์ให้เป็นแอสพาราจีน จะทำให้อัตราการถ่ายเทอิเล็กตรอนระหว่างโดเมนย่อย FMN กับฮีมในโดเมนออกซิจีเนสลดลง โดยค่าคงที่อัตราลดลงประมาณหนึ่งในสามจากการลดลงของความสามารถในการจับตัวระหว่างโดเมน ซึ่งได้รับการยืนยันด้วยเทคนิคเรโซแนนซ์พาราแมกเนติกอิเล็กตรอน (EPR) ในบริเวณจับ H4B ของโดเมนออกซิจีเนส กรดอะมิโนตำแหน่ง Arg375, Trp455, Trp457 และ Phe470 มีความสำคัญอย่างยิ่งต่อการจับโคแฟกเตอร์ H4B และการรวมตัวเป็นไดเมอร์และการทำงานของเอนไซม์ในเวลาต่อมา นอกจากนี้ Arg375 ยังเกี่ยวข้องกับการปรับหน้าที่รีดอกซ์ของ H4B อีกด้วย
ตำแหน่ง Phe831 และ Leu832 ในโดเมนรีดักเทสเป็นส่วนสำคัญของแกนไม่ชอบน้ำ (hydrophobic core) เมื่อกรดอะมิโนทั้งสองถูกแทนที่ด้วยกรดอะมิโนที่มีขั้ว (เซรีนและโพรลีนตามลำดับ) จะทำให้กิจกรรมของ iNOS ลดลงอย่างมาก ไม่น่าแปลกใจที่การกลายพันธุ์ของ Cys194 ซึ่งเป็นตำแหน่งจับฮีมให้กลายเป็นอะลานีน ส่งผลให้เอนไซม์ไม่สามารถจับฮีมได้ และยิ่งไปกว่านั้น เอนไซม์ยังไม่สามารถรวมตัวเป็นไดเมอร์ได้อีกด้วย เนื่องจากปฏิสัมพันธ์ระหว่างหมู่โพรพิโอเนตของฮีมกับ H4B เป็นที่ทราบกันดี จึงเป็นไปได้ว่ามอนอเมอร์ที่ขาดฮีมก็จะขาด H4B ไปด้วย ซึ่ง H4B นี้มีความสำคัญต่อกระบวนการรวมตัวเป็นไดเมอร์ ส่วนการจับตัวของ CaM จะถูกรบกวนความเสถียรจากการกลายพันธุ์ของ Ser562 และ Cys563 ใน iNOS ผ่านการทำลายเครือข่ายพันธะไฮโดรเจนที่สำคัญระหว่าง CaM กับ iNOS
หน้าที่ทางชีวภาพของ NO ที่ผลิตโดย iNOS
บทบาทหลักของ iNOS ในสรีรวิทยาของมนุษย์คือการทำลายเชื้อโรคที่บุกรุกเข้ามา ดังที่กล่าวไปแล้วว่า iNOS และ NO ถูกผลิตขึ้นเป็นส่วนหนึ่งของการตอบสนองทางภูมิคุ้มกันและมีบทบาทในการควบคุม เนื่องจาก NO สามารถแพร่กระจายได้ง่าย จึงมีชะตากรรมที่หลากหลาย ความสัมพันธ์อันสูงของ NO กับธาตุเหล็กทำให้มันสามารถทำลายหรือทำให้เอนไซม์ที่มีฮีมเป็นองค์ประกอบหรือกลุ่มเหล็ก-ซัลเฟอร์หยุดทำงานได้ NO อาจก่อให้เกิดปฏิกิริยาไนโตรซิเลชันของซีสเทอีน (cysteine nitrosylation) หรือรวมตัวกับซุปเปอร์ออกไซด์เพื่อสร้างเปอร์ออกซีไนไตรต์ (ONOO⁻) ซึ่งเป็นสารออกซิแดนท์และสารไนเตรตที่มีฤทธิ์รุนแรง ทั้ง NO และ ONOO⁻ สามารถก่อให้เกิดความเสียหายต่อดีเอ็นเอผ่านกระบวนการดีอะมิเนชันเชิงออกซิเดชัน (oxidative deamination) ด้วยอายุครึ่งชีวิตทางชีวภาพที่สั้นของ NO และ ONOO⁻ กลไกการทำลายเหล่านี้จึงมุ่งเป้าไปที่เชื้อโรคที่บุกรุกเป็นหลัก ช่วยหลีกเลี่ยงความเป็นพิษต่อระบบร่างกายโดยรวม
สุดท้ายนี้ การผลิตไซคลิกกัวโนซีนโมโนฟอสเฟต (cGMP) จะเพิ่มขึ้นผ่านการกระตุ้นเอนไซม์กัวนิเลตไซเคลส (guanylate cyclase) โดย NO cGMP เป็นสารสื่อสัญญาณลำดับที่สองชนิดละลายน้ำได้ ซึ่งมีบทบาทในกระบวนการส่งสัญญาณปลายน้ำหลากหลายรูปแบบ ผ่านการกระตุ้นโปรตีนไคเนสชนิดต่างๆ ช่องไอออน และฟอสโฟไดเอสเทอเรส
การควบคุมการทำงานของ iNOS
กิจกรรมของ iNOS ที่ถูกแสดงออกแล้วนั้นถูกควบคุมโดยปัจจัยหลายประการ ได้แก่ ปริมาณของสารตั้งต้นและโคแฟกเตอร์ ปฏิสัมพันธ์กับโปรตีนอื่นๆ และการหยุดทำงานของตัวเอง
ปริมาณของสารตั้งต้นและโคแฟกเตอร์
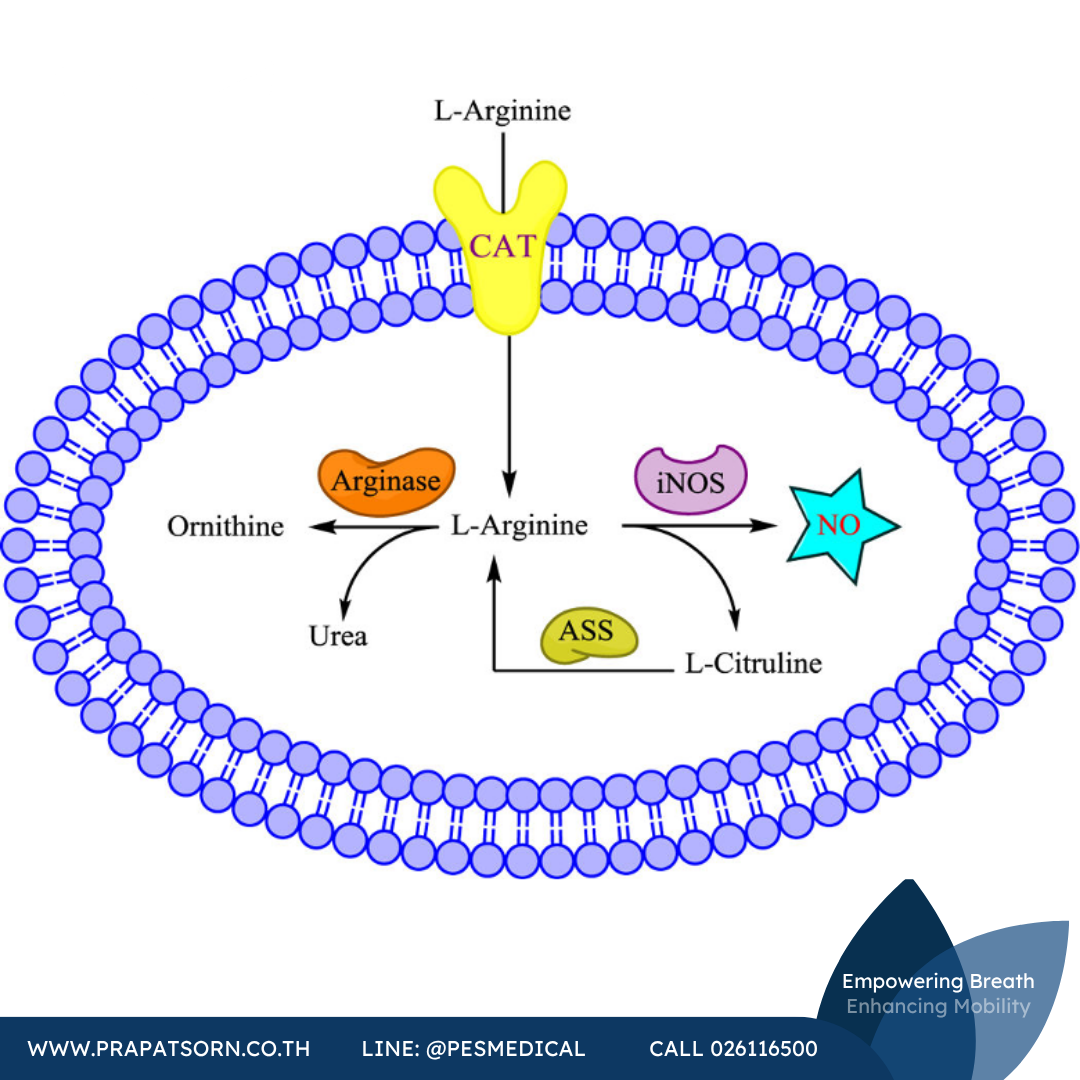
แอล-อาร์จินีนซึ่งเป็นสารตั้งต้นของ iNOS ยังมีความจำเป็นต่อการสังเคราะห์โปรตีนและกรดอะมิโนอื่นๆ และช่วยกำจัดแอมโมเนียที่เป็นพิษออกจากร่างกายในวัฏจักรยูเรีย (urea cycle) ในวัฏจักรยูเรียนี้ เอนไซม์อาร์จิเนส (arginase) จะเปลี่ยนอาร์จินีนให้เป็นออร์นิทีนและยูเรีย ซึ่งส่งผลต่อกิจกรรมของ iNOS ผ่านการแย่งชิงสารตั้งต้นชนิดเดียวกัน อันที่จริงมีการพบว่าการยับยั้งอาร์จิเนสช่วยเพิ่มการสังเคราะห์ NO ในแมคโครฟาจของกระต่ายและหนู และเพิ่มปริมาณโปรตีนที่ถูกไนเตรตและเอส-ไนโตรซิเลตในปอดหนูที่มีการอักเสบ ซึ่งอาจเป็นผลจากการเสริมการผลิต NO ในทางกลับกัน การแสดงออกมากเกินไปของอาร์จิเนสสามารถยับยั้งกิจกรรมของ iNOS และส่งเสริมการตอบสนองต่อการอักเสบที่ผิดปกติ ตัวอย่างเช่น การแสดงออกมากเกินไปของอาร์จิเนสในเซลล์เคอราติโนไซต์ของมนุษย์มีความเกี่ยวข้องกับกลไกการเกิดโรคสะเก็ดเงิน (psoriasis) ซึ่งเป็นโรคผิวหนังอักเสบ อาจเป็นผลมาจากการลดลงของกิจกรรม iNOS
ปริมาณของแอล-อาร์จินีนยังถูกควบคุมโดยตัวขนส่งกรดอะมิโนชนิดประจุบวก ได้แก่ CAT1, CAT2 และ CAT3 ซึ่งทำหน้าที่ขนส่งแอล-อาร์จินีนเข้าสู่เซลล์ สภาวะที่ใช้เหนี่ยวนำ iNOS ยังช่วยเพิ่มการแสดงออกของตัวขนส่งอาร์จินีนเหล่านี้ ทำให้การดูดซึมอาร์จินีนเพิ่มขึ้นตามไปด้วย ในแมคโครฟาจของสัตว์ฟันแทะ การเหนี่ยวนำ iNOS ด้วย LPS มาพร้อมกับการเพิ่มขึ้นของ CAT และการดูดซึมแอล-อาร์จินีนที่มากขึ้น ในทำนองเดียวกัน การขาดตัวขนส่งเหล่านี้จะลดกิจกรรมของ iNOS ลง การลบยีนตัวขนส่งอาร์จินีน CAT2 ทำให้กิจกรรมของ iNOS ในแอสโทรไซต์ที่ถูกกระตุ้นลดลงถึงร้อยละ 84 และยังลดการสังเคราะห์ NO ในแมคโครฟาจอีกด้วย
นอกจากนี้ แอล-ซิทรูลลีนซึ่งเป็นผลพลอยได้จากการผลิต NO สามารถถูกเปลี่ยนกลับเป็นอาร์จินีนได้ผ่านเอนไซม์อาร์จิโนซักซิเนตซินเทส (arginosuccinate synthase) โดย iNOS และเอนไซม์นี้จะถูกเหนี่ยวนำให้แสดงออกพร้อมกันในแมคโครฟาจ เซลล์กล้ามเนื้อเรียบของหลอดเลือด ไมโครเกลีย และเซลล์ประสาท ความสามารถของเซลล์ในการสร้างอาร์จินีนกลับคืนจากซิทรูลลีนนี้มีความจำเป็นเพื่อให้มีอาร์จินีนเพียงพอสำหรับ iNOS ในการรักษาบทบาทด้านสภาวะธำรงดุล (homeostasis)
ความพร้อมของโคแฟกเตอร์ที่จำเป็นอย่าง H4B ก็อาจส่งผลต่อกิจกรรมของ iNOS เช่นกัน H4B มีบทบาทสำคัญในการรวมตัวเป็นไดเมอร์ของ iNOS เพื่อให้กลายเป็นเอนไซม์ที่มีฤทธิ์ สายเซลล์ที่ผลิต H4B ได้น้อยจะมีกิจกรรม iNOS ต่ำ แต่การเสริม H4B จะช่วยเพิ่มการรวมตัวเป็นไดเมอร์ของ iNOS ได้ สารตั้งต้นของ H4B อย่างเซปิแอปเทอริน (sepiapterin) ก็สามารถช่วยฟื้นฟูกิจกรรมของ iNOS ได้เช่นกัน ที่น่าสนใจคือในระหว่างการอักเสบที่เกี่ยวข้องกับการหายของแผล iNOS จะแสดงออกและอยู่ร่วมตำแหน่งกับ GTP-cyclohydrolase I ซึ่งเป็นเอนไซม์สำคัญในวิถีการสังเคราะห์ H4B
ปฏิสัมพันธ์ระหว่างโปรตีน
มีโปรตีนหลายชนิดที่มีปฏิสัมพันธ์กับ iNOS และควบคุมกิจกรรมของมัน ในระบบประสาทส่วนกลาง พบว่า iNOS จับกับโปรตีนในไซโตซอลที่ชื่อว่าคาลิริน (kalirin โดยเฉพาะคาลิริน-7) ในสมองของหนูที่ได้รับ LPS การศึกษาพบว่าคาลิรินมีปฏิสัมพันธ์กับ iNOS ในรูปมอนอเมอร์แต่ไม่ใช่ไดเมอร์ ในเซลล์ที่แสดงออกคาลิริน iNOS ส่วนใหญ่จะอยู่ในรูปเฮเทอโรไดเมอร์ร่วมกับคาลิริน โดยมี iNOS แบบมอนอเมอร์เหลืออยู่น้อยมาก ซึ่งบ่งชี้ว่าคาลิรินยับยั้งกิจกรรมของ iNOS โดยการป้องกันการรวมตัวเป็นโฮโมไดเมอร์ของ iNOS การแสดงออกของคาลิรินในระดับต่ำมีความสัมพันธ์กับกิจกรรม iNOS ที่เพิ่มขึ้นในฮิปโปแคมปัสของผู้ป่วยอัลไซเมอร์ เนื่องจาก NO ส่วนเกินที่ผลิตขึ้นระหว่างการอักเสบเป็นพิษต่อเซลล์ประสาท คาลิรินจึงอาจออกฤทธิ์ปกป้องเซลล์ประสาทด้วยการลดการทำงานของ iNOS ลง
ในแมคโครฟาจของหนู iNOS มีปฏิสัมพันธ์กับโปรตีนขนาด 110 กิโลดาลตันที่เรียกว่า NAP110 (NOS-associated protein 110) สารประกอบเชิงซ้อนของ NAP110 และ iNOS พบได้ในม้ามและแมคโครฟาจในช่องท้องของหนูที่ได้รับ IFN-γ และ LPS การแสดงออกร่วมกันของ NAP110 และ iNOS ลดกิจกรรมของ iNOS ลงถึงร้อยละ 90 แม้ปริมาณโปรตีน iNOS จะไม่เปลี่ยนแปลงก็ตาม เช่นเดียวกับคาลิริน NAP110 ยับยั้งกิจกรรมของ iNOS โดยการมีปฏิสัมพันธ์กับ iNOS แบบมอนอเมอร์และป้องกันการรวมตัวเป็นไดเมอร์ของเอนไซม์ ดังนั้น NAP110 จึงอาจทำหน้าที่เป็นสวิตช์ควบคุมเพื่อป้องกันไม่ให้ระดับ NO สูงจนเป็นพิษ
นอกจากนี้ iNOS ยังพบว่ามีปฏิสัมพันธ์โดยตรงกับ Rho GTPase Rac2 โดยส่วนใหญ่ผ่านทางโดเมนออกซิจีเนส การแสดงออกมากเกินไปของ Rac2 ในเซลล์ RAW 264.7 ช่วยเสริมฤทธิ์กิจกรรม iNOS ที่ถูกกระตุ้นด้วย LPS อีกการศึกษาหนึ่งแสดงให้เห็นว่า iNOS และ Rac2 มีปฏิสัมพันธ์กันในนิวโทรฟิลของมนุษย์ ปฏิสัมพันธ์นี้อาจมีบทบาทสำคัญในการป้องกันเชื้อจุลชีพ เนื่องจากเซลล์ที่ได้รับสารยับยั้ง Rac2 สร้างซุปเปอร์ออกไซด์ (ซึ่งทำปฏิกิริยากับ NO เพื่อผลิต ONOO⁻ ที่เป็นพิษ) ได้น้อยลง ผลการศึกษานี้ชี้ให้เห็นว่า Rac2 สามารถเสริมการสร้าง NO ผ่าน iNOS ได้ ในขณะที่ Yoshida และ Xia พบว่าโปรตีนชาเปอโรน heat shock protein 90 (hsp90) ก็สามารถเพิ่มกิจกรรมของ iNOS ได้ในลักษณะขึ้นกับขนาดยา และโปรตีนทั้งสองชนิดสามารถตกตะกอนร่วมกันได้จากแมคโครฟาจที่ถูกกระตุ้น
การหยุดทำงานของตัวเอง (Auto-inactivation)
NO เองก็อาจมีบทบาทในการควบคุมกิจกรรมของ iNOS ในทางลบได้เช่นกัน ในแมคโครฟาจของหนูที่ถูกกระตุ้น กิจกรรมของ iNOS จะเพิ่มขึ้นเมื่อมีการเติมฮีโมโกลบิน ซึ่งเป็นตัวจับ NO (NO scavenger) การเติมตัวให้ NO อย่าง S-nitrosoacetylpenicillamine หรือ S-nitrosoglutathione จะยับยั้งกิจกรรมของ iNOS อย่างมีนัยสำคัญ ซึ่งบ่งชี้ว่า NO อาจเกี่ยวข้องกับกลไกป้อนกลับเชิงลบ (negative feedback) บางอย่าง ยิ่งไปกว่านั้น กิจกรรมของ iNOS ไม่สามารถฟื้นคืนได้ในเซลล์ที่ถูกกระตุ้นแม้จะนำตัวให้ NO ออกไปแล้ว ซึ่งเป็นข้อบ่งชี้ถึงการจับตัวแบบไม่ผันกลับของ NO กับ iNOS การศึกษาจลนศาสตร์แสดงให้เห็นว่ากลไกหลักของการหยุดทำงานของตัวเองใน iNOS เกิดผ่านปฏิกิริยาเอส-ไนโตรเซชันของกลุ่มซิงก์เททระไทโอเลต ซึ่งทำให้เกิดการสูญเสียสังกะสี การแยกตัวของไดเมอร์ iNOS แบบไม่ผันกลับ และการสูญเสียกิจกรรมตามมา สารรีดิวซ์สามารถปกป้อง iNOS จากการหยุดทำงานนี้ได้ ซึ่งบ่งชี้ว่าเอส-ไนโตรเซชันเป็นวิถีการหยุดทำงานหลักอย่างแท้จริง
การแสดงออกของยีนและวิถีสัญญาณของ iNOS
การแสดงออกของ iNOS ถูกควบคุมโดยหลักผ่านกลไกการถอดรหัส (transcriptional regulation) ซึ่งอาจแตกต่างกันไปตามชนิดของเซลล์หรือชนิดของสิ่งมีชีวิต โดยทั่วไป ไซโตไคน์ที่ก่อการอักเสบ เช่น tumor necrosis factor alpha (TNF-α), interleukin-1 (IL-1), interferon-γ (IFN-γ) และ LPS จะจับกับตัวรับบนผิวเซลล์ก่อน แล้วกระตุ้นเอนไซม์ไคเนส ซึ่งนำไปสู่การฟอสโฟรีเลชันของโปรตีนภายในเซลล์หลายชนิด และการกระตุ้นปัจจัยการถอดรหัสจำเพาะตามมา รวมถึงปัจจัยการถอดรหัสตระกูล nuclear factor κB (NF-κB) เช่น signal transducer and activator of transcription 1α (STAT-1α) ปัจจัยที่ถูกกระตุ้นเหล่านี้จะเคลื่อนย้ายเข้าสู่นิวเคลียส ซึ่งจะจับกับบริเวณโปรโมเตอร์ของยีน iNOS และกระตุ้นการแสดงออกของ iNOS
ตัวกระตุ้นที่ต่างกันจะเหนี่ยวนำวิถีสัญญาณที่ต่างกัน วิถีหลักวิถีหนึ่งเกี่ยวข้องกับการกระตุ้น NF-κB ซึ่งเป็นเป้าหมายสำคัญของ LPS, IL-1β และ TNF ในแมคโครฟาจ LPS จะกระตุ้นตัวรับแบบ toll-like receptor (TLR4) บนผิวเซลล์ก่อน สารประกอบเชิงซ้อน TLR4 ประกอบด้วยโปรตีนหลายชนิด ได้แก่ TNF-receptor associated factor 6 (TRAF6), myeloid differentiation primary response 88 (MyD88) และเอนไซม์ไคเนสชนิดหนึ่ง เนื่องจาก NF-κB ในแมคโครฟาจอยู่ในรูปไดเมอร์และโดยปกติจะถูกทำให้ไม่ทำงานและถูกจับไว้โดยตัวยับยั้ง κB (IκB) การกระตุ้นวิถีนี้จะเหนี่ยวนำให้เกิดปฏิกิริยาลูกโซ่ของไคเนส (หนึ่งในนั้นผ่านทาง mitogen-activated protein kinase kinase หรือ MEKK) ซึ่งนำไปสู่การฟอสโฟรีเลชันของ inhibitor of κB kinase (IKK) และการฟอสโฟรีเลชันของ IκB ตามมา เมื่อถูกฟอสโฟรีเลตแล้ว IκB จะถูกยูบิควิทิเนตและถูกย่อยสลายในโปรตีเอโซม ทำให้ NF-κB ถูกปลดปล่อยออกมา จากนั้น NF-κB จะเคลื่อนย้ายเข้านิวเคลียส มีปฏิสัมพันธ์กับโปรโมเตอร์ของ iNOS และกระตุ้นการถอดรหัสของยีน iNOS
ในทางตรงกันข้าม IFN-γ จะกระตุ้นวิถี JAK/STAT-1α กล่าวโดยสรุป การกระตุ้นเซลล์ด้วย IFN-γ จะทำให้เกิดการรวมตัวเป็นไดเมอร์ของตัวรับ IFN-γ และการกระตุ้นเอนไซม์ Janus kinase (JAK เช่น JAK2) ซึ่งจะฟอสโฟรีเลต STAT-1α จากนั้น STAT-1α เองก็จะรวมตัวเป็นไดเมอร์และเคลื่อนย้ายเข้าสู่นิวเคลียส ซึ่งจะส่งเสริมการสังเคราะห์ interferon regulatory factor 1 (IRF-1) จากนั้น IRF-1 จะจับกับโปรโมเตอร์ของยีน iNOS และกระตุ้นการแสดงออกของ iNOS ต่อไป
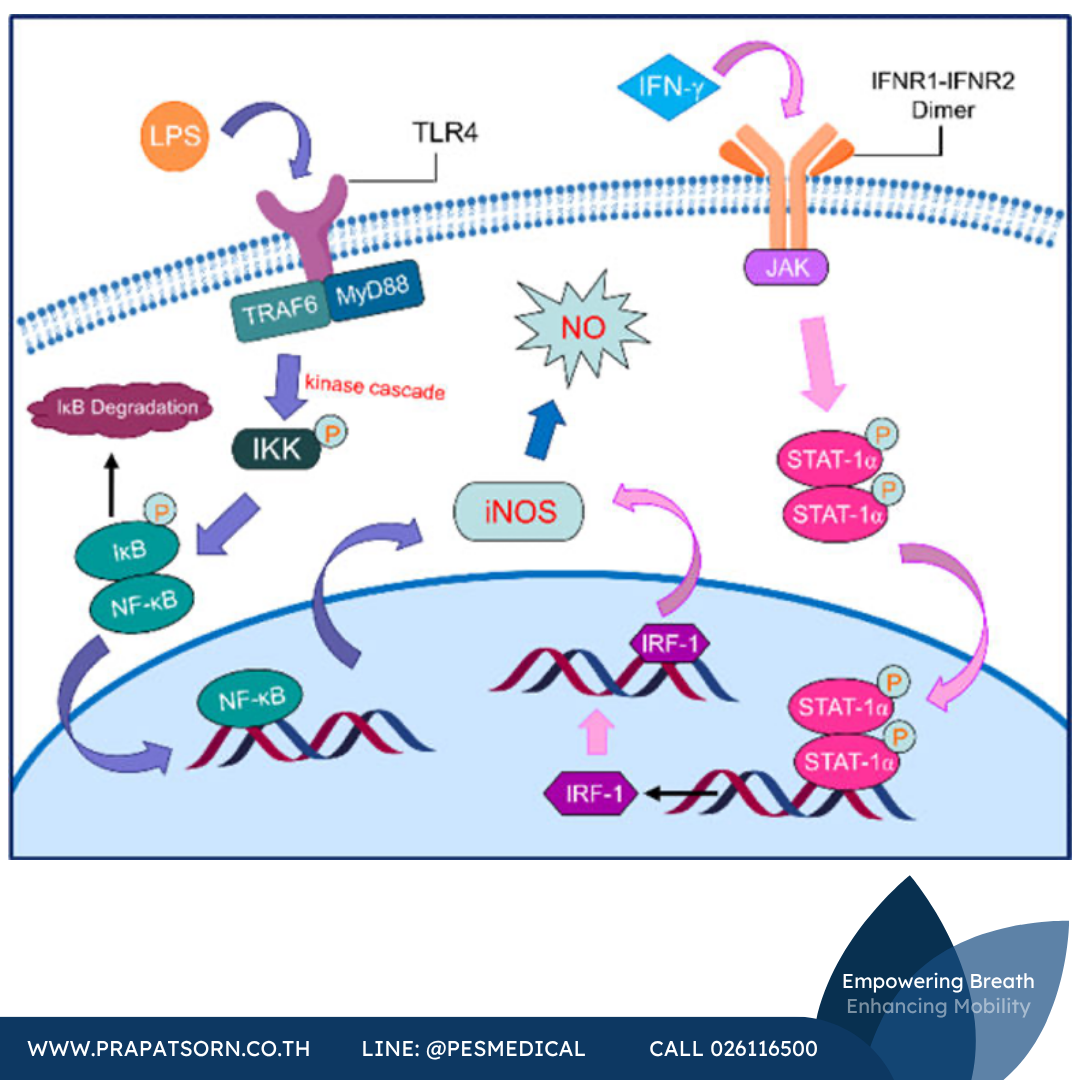
ภาพที่ 4: วิถีสัญญาณที่แตกต่างกันซึ่งกระตุ้นโดยตัวเหนี่ยวนำ iNOS ชนิดต่างๆ (ซ้าย: วิถี NF-κB, ขวา: วิถี JAK/STAT-1α)
การกระจายตัวและการเหนี่ยวนำให้เกิด iNOS
iNOS ถูกทำให้บริสุทธิ์ครั้งแรกจากแมคโครฟาจของหนู หลังจากนั้นมีการค้นพบว่าเซลล์หลายชนิดจากสิ่งมีชีวิตต่างสายพันธุ์สามารถถูกเหนี่ยวนำให้แสดงออก iNOS ได้เมื่อถูกกระตุ้น ทั้งเซลล์ตับ เซลล์กล้ามเนื้อเรียบ คอนโดรไซต์ เซลล์เกลีย แอสโทรไซต์ เซลล์ประสาท และกล้ามเนื้อหัวใจ การโคลนสายดีเอ็นเอเสริม (cDNA) ของ iNOS ที่มีความยาวเต็มโมเลกุลจากแมคโครฟาจของหนู RAW 264.7 ได้สำเร็จ ทำให้ทราบลำดับกรดอะมิโนของโปรตีนและเข้าใจโครงสร้างและการควบคุมของมันมากขึ้น ลำดับกรดอะมิโนของ iNOS จากแมคโครฟาจหนูมีความคล้ายคลึงกับ nNOS ของหนูแรทถึงร้อยละ 51
เมื่อเปรียบเทียบกับสิ่งมีชีวิตชนิดอื่น การเหนี่ยวนำ iNOS ในเซลล์ของมนุษย์นั้นมีข้อจำกัดมากกว่า แมคโครฟาจของมนุษย์ไม่สามารถผลิต NO ได้เพียงพอภายใต้เงื่อนไขการเหนี่ยวนำหลายรูปแบบ ซึ่งอาจเป็นผลจากการสังเคราะห์ H4B ที่จำกัด ซึ่งเป็นโคแฟกเตอร์สำคัญของ iNOS ในแมคโครฟาจของมนุษย์ การเหนี่ยวนำ iNOS ในมนุษย์ที่สำเร็จเป็นครั้งแรกเกิดขึ้นในเซลล์ตับ cDNA ของ iNOS จากตับมนุษย์มีความคล้ายคลึงของลำดับกับ iNOS จากแมคโครฟาจหนูถึงร้อยละ 80 การเหนี่ยวนำ iNOS ของมนุษย์ยังประสบความสำเร็จในคอนโดรไซต์ของมนุษย์โดยใช้ IL-1β เช่นกัน โดย cDNA ที่โคลนจากเซลล์ชนิดนี้มีความคล้ายคลึงกับ iNOS จากแมคโครฟาจหนูถึงร้อยละ 88 ซึ่งบ่งชี้ว่าโครงสร้างและหน้าที่ของ iNOS ยังคงได้รับการอนุรักษ์ไว้ในสิ่งมีชีวิตและชนิดเซลล์ที่ต่างกัน
ในระบบประสาทส่วนกลาง iNOS ถูกแสดงออกในเซลล์เกลียหลายชนิด รวมทั้งแอสโทรไซต์และไมโครเกลีย (ซึ่งได้รับฉายาว่า “แมคโครฟาจของสมอง”) เพื่อตอบสนองต่อสิ่งกระตุ้นต่างๆ เมื่อเทียบกับเซลล์เกลียแล้ว การแสดงออกของ iNOS ในเซลล์ประสาทค่อนข้างจำกัดมาก แต่ก็ยังมีหลักฐานยืนยันการแสดงออกในเซลล์เหล่านี้อยู่บ้าง ตัวอย่างเช่น การรักษาสายเซลล์นิวโรบลาสโตมาของมนุษย์หลายสายพันธุ์ (ซึ่งใช้เป็นแบบจำลองของเซลล์ประสาทมนุษย์อย่างแพร่หลาย) ด้วยไซโตไคน์ ส่งผลให้เกิดการแสดงออกของยีน iNOS การแสดงออกของ iNOS ในเซลล์ประสาทอาจเกิดขึ้นได้ในโรคอัลไซเมอร์ หรือภายหลังภาวะพร่องออกซิเจนหรือภาวะขาดเลือด
TNF, IL-1β, IFN-γ และ LPS เป็นตัวกระตุ้นหลักของการแสดงออก iNOS การผสมผสานของตัวกระตุ้นเหล่านี้สามารถก่อให้เกิดผลเสริมฤทธิ์กัน (synergistic effects) ในเซลล์บางชนิด และบางครั้งจำเป็นต้องใช้ร่วมกันเพื่อกระตุ้นการผลิต NO โดยเฉพาะในเซลล์ที่ไม่ใช่เซลล์ภูมิคุ้มกัน ซึ่ง LPS เพียงอย่างเดียวจะกระตุ้น iNOS ได้เพียงเล็กน้อย ตัวอย่างเช่น ในไมโอไซต์กล้ามเนื้อโครงร่างของหนู ไม่มีตัวกระตุ้นใดตัวหนึ่งที่สามารถกระตุ้นการแสดงออกของ iNOS ได้เพียงลำพัง แต่การกระตุ้นเซลล์ด้วยทั้ง IFN-γ และ TNF หรือ IL-1 (หรือทั้งสามไซโตไคน์ร่วมกัน) ส่งผลให้เกิดการผลิต NO อย่างมีนัยสำคัญ สิ่งกระตุ้นและเงื่อนไขที่กำหนดการแสดงออกของ iNOS ขึ้นอยู่กับชนิดของเซลล์และสิ่งมีชีวิต ตัวอย่างเช่น LPS เพียงอย่างเดียวสามารถกระตุ้นการแสดงออกของ iNOS ในเซลล์เกลียของหนูแรทและหนูเมาส์ได้ แต่ไม่สามารถกระตุ้นในเซลล์เกลียของมนุษย์ได้
นอกเหนือจาก LPS และไซโตไคน์แล้ว ไวรัสที่บุกรุกเข้าสู่ร่างกายก็สามารถกระตุ้นการเหนี่ยวนำ iNOS ในเซลล์ได้เช่นกัน เชื้อไวรัสภูมิคุ้มกันบกพร่องของมนุษย์ (HIV) กระตุ้นการแสดงออกของ iNOS ในเซลล์เกลียของมนุษย์ และการแสดงออกของ iNOS ในระดับสูงยังตรวจพบได้ในเซลล์ตับของผู้ป่วยที่ติดเชื้อไวรัสตับอักเสบบี (HBV) การถ่ายยีน HBV เข้าสู่เซลล์ตับส่งผลให้เกิดการแสดงออกของ iNOS ซึ่งบ่งชี้ว่าไวรัสสามารถเพิ่มการแสดงออกของ iNOS ในเซลล์ตับของมนุษย์ได้
iNOS กับสภาวะโรคต่างๆ
แม้ iNOS จะมีความจำเป็นต่อสรีรวิทยาปกติของร่างกาย แต่ปริมาณ NO ที่มากเกินไปอันเป็นผลจากการแสดงออกมากเกินไปหรือการควบคุมที่ผิดปกติของ iNOS ก็มีความเกี่ยวข้องกับโรคของมนุษย์หลายชนิด มีการศึกษาจำนวนมากแสดงให้เห็นว่า NO ส่วนเกินมีบทบาทในภาวะติดเชื้อในกระแสเลือด (sepsis) ในแบบจำลองสัตว์ทดลอง การผลิต NO ในระดับสูงมีส่วนทำให้เกิดการขยายหลอดเลือด ความดันโลหิตต่ำ และความผิดปกติของระบบหัวใจและหลอดเลือดในระหว่างภาวะเอนโดทอกซีเมียและภาวะช็อกที่เกิดจากไซโตไคน์ ระดับไนไตรต์และไนเตรตที่สูง (ผลิตภัณฑ์ที่เสถียรสองชนิดจากการผลิต NO) พบได้ในพลาสมาของผู้ป่วยที่มีภาวะช็อกจากการติดเชื้อ และยังตรวจพบกิจกรรมของ iNOS ในเนื้อเยื่อเน่าตายของผู้ป่วยภาวะติดเชื้อในกระแสเลือด ทั้งในไขมัน กล้ามเนื้อ และหลอดเลือดแดง (แต่ไม่พบในผิวหนัง) นอกจากนี้ยังพบกิจกรรมของ iNOS ที่เพิ่มขึ้นในแมคโครฟาจปอดของผู้ป่วยกลุ่มอาการหายใจลำบากเฉียบพลัน (ARDS) ภายหลังภาวะติดเชื้อในกระแสเลือด ด้วยความรุนแรงถึงชีวิตของภาวะช็อกจากการติดเชื้อ ข้อบ่งชี้นี้จึงเป็นแรงผลักดันสำคัญให้เกิดการพัฒนาสารยับยั้ง iNOS จำนวนมาก
การแสดงออกของ iNOS ในระดับสูงยังพบได้ในผู้ป่วยที่ติดเชื้อก่อโรคชนิดต่างๆ โดยเฉพาะเชื้อ HIV วัณโรค (Mycobacterium tuberculosis) และเชื้อมาลาเรีย (Plasmodium falciparum) ทั้งการติดเชื้อ HIV และวัณโรคต่างกระตุ้นการแสดงออกของ iNOS ในเซลล์ที่มาจากสมอง และในกรณีของ HIV นั้น NO ที่เกิดขึ้นอาจมีส่วนทำให้เกิดความเสียหายของเซลล์ประสาทในภาวะสมองเสื่อมที่สัมพันธ์กับโรคเอดส์ นอกจากนี้ การเหนี่ยวนำ iNOS ยังอาจเชื่อมโยงกับภาวะดื้ออินซูลินที่สัมพันธ์กับเบาหวานและโรคอ้วน เนื่องจากเป็นที่ทราบกันว่า iNOS ถูกแสดงออกทั่วเนื้อเยื่อที่ไวต่ออินซูลิน และโรคอ้วนเพิ่มระดับ iNOS ในกล้ามเนื้อ เนื้อเยื่อไขมัน และหลอดเลือด การศึกษาในแบบจำลองหนูแรทแสดงให้เห็นว่าเซลล์เกาะตับอ่อนของหนูมีความไวต่อพิษของ NO อย่างมาก และพบการแสดงออกมากเกินไปของ iNOS ในแมคโครฟาจของตับอ่อนในหนูแรทที่มีภาวะก่อนเบาหวาน บทบาทของ iNOS ในภาวะดื้ออินซูลินอาจเกิดขึ้นบางส่วนผ่านกระบวนการเอส-ไนโตรซิเลชันหรือไทโรซีนไนเตรชันของโปรตีนในวิถีสัญญาณอินซูลิน
การแสดงออกของ iNOS ในระดับสูงยังพบได้ในเนื้องอกมะเร็งหลายชนิด ทั้งมะเร็งลำไส้ใหญ่ กระเพาะปัสสาวะ เต้านม และปอด บทบาทของ iNOS ในมะเร็งค่อนข้างซับซ้อน เนื่องจากมีทั้งฤทธิ์ส่งเสริมเนื้องอก (tumor promoter) และฤทธิ์ยับยั้งเนื้องอก (tumor suppressor) ขึ้นอยู่กับชนิดของเนื้องอกและสภาพแวดล้อมเฉพาะที่ ปฏิกิริยาไนโตรเซชันหรือไนโตรซิเลชัน (จาก NO และผลิตภัณฑ์ของมัน) ในเซลล์มะเร็งสามารถส่งเสริมการเพิ่มจำนวนเซลล์ การแพร่กระจาย และแม้แต่การดื้อยา เมื่อไม่นานมานี้ พบว่าการแสดงออกของ iNOS ในระดับสูงมีความสัมพันธ์กับอัตราการรอดชีวิตที่ต่ำในผู้ป่วยมะเร็งเต้านมชนิดสามลบ (triple-negative breast cancer) นอกจากนี้ NO ที่ผลิตโดย iNOS ภายในเนื้องอกเองยังสามารถยับยั้งการเพิ่มจำนวนของทีลิมโฟไซต์ (ดังที่พบในแบบจำลองมะเร็งลำไส้ใหญ่ของหนู) ซึ่งอาจอธิบายได้ว่าเหตุใดเนื้องอกจึงสามารถกดการทำงานของระบบภูมิคุ้มกันของโฮสต์ได้
การศึกษาต่างๆ ยังบ่งชี้ว่า iNOS (ทั้งที่แสดงออกในระบบประสาทส่วนกลางและส่วนปลาย) อาจมีบทบาทในการเกิดและการรับรู้ความเจ็บปวดจากการอักเสบและความเจ็บปวดทางระบบประสาท Meller และคณะพบว่า LPS และไซโตไคน์ทำให้เกิดภาวะไวต่อความปวดจากความร้อน (thermal hyperalgesia) ซึ่งอาจเกิดผ่านการเหนี่ยวนำ iNOS ในเซลล์เกลียของไขสันหลัง ในทำนองเดียวกัน ในแบบจำลองสัตว์ทดลองของความเจ็บปวดทางระบบประสาท การเพิ่มขึ้นของ iNOS มีส่วนทำให้เกิดปฏิกิริยาอักเสบเฉพาะที่ เมื่อไม่นานมานี้ พบว่าแม้แต่ความเจ็บปวดที่สัมพันธ์กับมะเร็งก็มีความเกี่ยวข้องกับการเพิ่มขึ้นของ iNOS (และ nNOS) ในหนูที่เป็นมะเร็งกระดูก ด้วยเหตุนี้ การรักษาความเจ็บปวดจึงเป็นแรงผลักดันสำคัญอีกประการหนึ่งสำหรับการพัฒนายาที่ยับยั้ง iNOS
สมองปกติดูเหมือนจะไม่แสดงออก iNOS มากนัก แต่ภายหลังการบาดเจ็บ ความเสียหายหรือสิ่งกระตุ้นจากการอักเสบ หรือภาวะพร่องออกซิเจน (เช่นจากโรคหลอดเลือดสมองหรือภาวะขาดเลือด) การแสดงออกของ iNOS ในเซลล์เกลียและเซลล์ประสาทที่ถูกกระตุ้นสามารถเกิดขึ้นได้ เซลล์ที่ผลิต iNOS เหล่านี้บางส่วนสะสมตัวอยู่รอบบริเวณที่เสียหายในสมอง ซึ่งอาจบ่งชี้ถึงความเชื่อมโยงเชิงการอักเสบระหว่าง NO ที่มาจากเซลล์เกลียกับการบาดเจ็บของเซลล์ประสาท พบ iNOS ในสมองของผู้ป่วยโรคอัลไซเมอร์ และยังพบระดับ iNOS ที่สูงในแบบจำลองสัตว์ทดลองหลายรูปแบบของโรคพาร์กินสัน อย่างไรก็ตาม หนูที่ถูกน็อกเอาต์ยีน iNOS และผสมพันธุ์ข้ามสายพันธุ์กับหนูดัดแปลงพันธุกรรมที่ผลิตแผ่นเบตา-อะไมลอยด์ กลับแสดงอาการคล้ายอัลไซเมอร์ในอัตราที่สูงกว่าหนูดัดแปลงพันธุกรรมที่ผลิตแผ่นเบตา-อะไมลอยด์เพียงอย่างเดียวมาก ซึ่งชี้ให้เห็นว่า iNOS อาจมีบทบาทปกป้อง และการยับยั้งมันอาจก่อให้เกิดผลเสียได้ ยังคงมีความไม่แน่นอนอยู่บ้างว่าการแสดงออกและกิจกรรมของ iNOS เป็นสาเหตุโดยตรงของความเสียหายของเซลล์ประสาท หรือเพียงแค่มีความสัมพันธ์กับบางระยะของโรคความเสื่อมของระบบประสาท (กล่าวคือ เป็นเพียงความสัมพันธ์ ไม่ใช่ความเป็นเหตุเป็นผล) หรืออาจมีฤทธิ์ปกป้องในบางกรณีด้วยซ้ำ
การพัฒนาสารยับยั้ง iNOS
วิธีการทดสอบกิจกรรมและการยับยั้ง iNOS
มีการทดสอบในหลอดทดลองหลายรูปแบบที่นิยมใช้ในการวัดกิจกรรมของ iNOS ระหว่างการพัฒนาสารยับยั้ง ระบบที่ใช้เอนไซม์บริสุทธิ์มักถูกใช้เป็นขั้นแรกเพื่อประเมินระดับการยับยั้ง iNOS วิธีหนึ่งที่นิยมใช้คือการวัดการเปลี่ยนแอล-อาร์จินีนที่ติดฉลากกัมมันตรังสี (3H หรือ 14C) ให้เป็นแอล-ซิทรูลลีนโดยใช้ iNOS ของมนุษย์หรือหนู แม้ว่าวิธีที่ใช้เอนไซม์แบบไม่ใช้สารกัมมันตรังสี เช่น การทดสอบด้วยฮีโมโกลบิน (hemoglobin capture assay ซึ่งวัดการออกซิไดซ์ฮีโมโกลบินเป็นเมทฮีโมโกลบินโดย NO ด้วยวิธีสเปกโทรโฟโตเมตรี) ก็ถูกนำมาใช้เช่นกัน ระบบที่ใช้เซลล์ (โดยเฉพาะสำหรับสารยับยั้งการรวมตัวเป็นไดเมอร์) ซึ่งใช้สายเซลล์แมคโครฟาจ (เช่น RAW 264.7) ถูกกระตุ้นให้แสดงออก iNOS ด้วยไซโตไคน์ ก็ถูกนำมาใช้เช่นกัน ในระบบเหล่านี้ การผลิต NO จะถูกติดตาม เช่น ผ่านการเติมรีเอเจนต์ Griess ลงในไลเสทของเซลล์ ซึ่งตรวจจับการผลิตไนไตรต์
การทดสอบในสัตว์ทดลอง (in vivo) ที่นิยมใช้มากที่สุดสำหรับการยับยั้ง iNOS คือแบบจำลองภาวะเอนโดทอกซีเมียในสัตว์ฟันแทะ ซึ่งใกล้เคียงกับลักษณะการอักเสบทั่วร่างกายที่พบในภาวะติดเชื้อในกระแสเลือด ในการทดสอบนี้ จะให้สารเอนโดทอกซิน (โดยปกติคือ LPS) แก่สัตว์ฟันแทะ ซึ่งก่อให้เกิดการอักเสบทั่วร่างกายพร้อมกับการเพิ่มขึ้นของไซโตไคน์ iNOS และการผลิต NO ซึ่งสามารถติดตามได้ผ่านระดับไนเตรต/ไนไตรต์ในพลาสมา
แบบจำลองฟอร์มาลินหรือคาราจีแนนมักถูกใช้เพื่อจำลองความเจ็บปวดจากการอักเสบ โดยจะฉีดสารละลายฟอร์มาลินปริมาณเล็กน้อยเข้าที่อุ้งเท้าของสัตว์ฟันแทะเพื่อก่อให้เกิดการอักเสบเฉพาะที่ แล้ววัดความถี่หรือระยะเวลาของพฤติกรรมแสดงความเจ็บปวด (การกระตุกหรือกัดและเลียอุ้งเท้า) ทั้งในกรณีที่มีและไม่มีสารยับยั้งที่สนใจ ในทำนองเดียวกัน การฉีดคาราจีแนนก่อให้เกิดอาการบวมน้ำ ความเจ็บปวด และภาวะไวต่อความปวดเฉพาะที่ เพื่อประเมินบทบาทของ iNOS ในความเจ็บปวดทางระบบประสาท จะใช้แบบจำลองการผ่าตัด เช่น แบบจำลอง Chung (การเหนี่ยวนำความเจ็บปวดผ่านการผูกเส้นประสาทไขสันหลังด้วยการผ่าตัด) หรือแบบจำลองการบาดเจ็บจากการกดทับเรื้อรัง (การผูกเส้นประสาทไซอาติก) เนื่องจากสารยับยั้ง iNOS ยังถูกศึกษาสำหรับโรคข้ออักเสบด้วย จึงมีการใช้แบบจำลองข้ออักเสบที่เหนี่ยวนำด้วยสารเสริมฤทธิ์ (adjuvant-induced arthritis model) เพื่อประเมินประสิทธิภาพ โดยจะให้สารเสริมฤทธิ์ (เช่น Freund’s Complete Adjuvant) แก่สัตว์ทดลองเพื่อเหนี่ยวนำให้เกิดลักษณะข้ออักเสบ จากนั้นจะสังเกตอาการข้ออักเสบทั้งในกรณีที่มีและไม่มียา และให้คะแนนทางคลินิก บางครั้งก็มีการชำแหละและตรวจสอบองค์ประกอบของข้อที่ได้รับผลกระทบภายหลังการเสียชีวิตด้วย
การยับยั้งแบบแข่งขันกับสารตั้งต้น (Competitive iNOS Inhibitors)
กลไกหลักของการยับยั้ง iNOS ที่ออกฤทธิ์โดยตรงต่อเอนไซม์มีอยู่สองรูปแบบ รูปแบบแรกคือสารยับยั้งส่วนใหญ่ที่ถูกค้นพบจะแข่งขันโดยตรงกับแอล-อาร์จินีน (มักออกฤทธิ์โดยการประสานพันธะกับกรดกลูตามิกที่ตำแหน่งเร่งปฏิกิริยา) โดยจะยับยั้งการจับตัวของแอล-อาร์จินีนแบบผันกลับได้ หรือแข่งขันกับแอล-อาร์จินีนแล้วทำให้เอนไซม์หยุดทำงานอย่างถาวรในลักษณะที่ขึ้นกับเวลา ความเข้มข้น หรือ NADPH เนื้อหาต่อไปนี้จะไล่เรียงพัฒนาการของสารยับยั้งกลุ่มนี้ตามลำดับเวลา
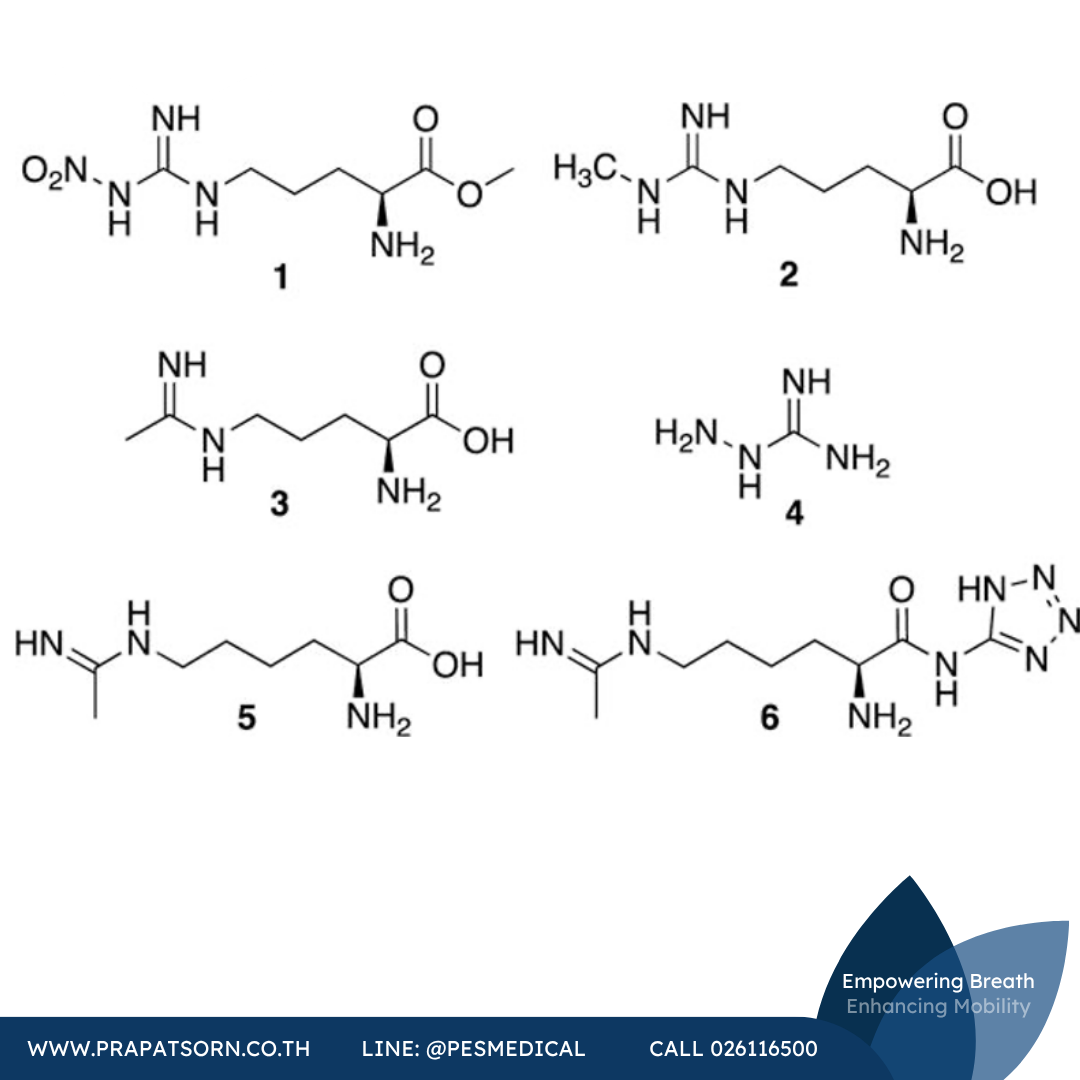
สารยับยั้ง iNOS แบบแข่งขันที่ถูกรายงานในยุคแรกๆ คือสารอนุพันธ์ของอาร์จินีน ได้แก่ แอล-ไนโตรอาร์จินีน สารเมทิลเอสเทอร์ที่มีชีวประสิทธิผลดีกว่าของมัน (L-NAME หรือสารประกอบ 1) และแอล-Nω-โมโนเมทิลอาร์จินีน (L-NMMA หรือสารประกอบ 2) แม้สารประกอบ 2 จะเป็นทั้งสารยับยั้งแบบแข่งขันและสารทำให้หยุดทำงานอย่างช้าๆ ของ iNOS (อาจผ่านสารมัธยันตร์ไฮดรอกซิเลตและการสูญเสียฮีม) แต่สารอนุพันธ์อาร์จินีนอย่างง่ายเหล่านี้กลับมีความจำเพาะต่อ iNOS น้อยมากเมื่อเทียบกับอีกสองไอโซฟอร์ม สารประกอบ 2 ยับยั้งทั้ง iNOS และ nNOS ส่วนสารประกอบ 1 (ในรูปกรดที่ผ่านการไฮโดรไลซ์ เนื่องจาก L-NAME เองเป็นเพียงสารยับยั้งฤทธิ์อ่อน) และสารประกอบ 2 กลับมีความจำเพาะต่อ eNOS มากกว่า ด้วยเหตุนี้ ทั้งไนโตรอาร์จินีนและสารประกอบ 2 จึงก่อให้เกิดความดันโลหิตสูงเมื่อให้แก่หนูแรทหรือมนุษย์ ซึ่งน่าจะเป็นผลจากการยับยั้งการขยายหลอดเลือดที่อาศัย eNOS
สารประกอบเอมิดีน N5-(1-iminoethyl)-L-ornithine (L-NIO หรือสารประกอบ 3) เป็นสารทำให้หยุดทำงานแบบแข่งขันที่อาศัยกลไกเฉพาะ (mechanism-based inactivator) โดยมีความจำเพาะต่อ iNOS ของหนูเมาส์มากกว่า nNOS ของหนูแรทประมาณ 1.8 เท่า แต่มีความจำเพาะต่อ eNOS ของวัวน้อยมาก ในแบบจำลองภาวะเอนโดทอกซีเมียของหนูแรท คณะวิจัยของ László ได้ศึกษาความสามารถของสารยับยั้ง iNOS ในการป้องกันความเสียหายของหลอดเลือดฝอยในลำไส้ใหญ่และการรั่วไหลของอัลบูมินที่เกิดจากเอนโดทอกซินอันเนื่องมาจาก NO ส่วนเกิน เมื่อให้ในช่วงที่ iNOS ถูกเหนี่ยวนำ (ประมาณ 4 ชั่วโมงหลังได้รับ LPS) สารประกอบ 3 ช่วยลดการรั่วไหลของลำไส้ใหญ่ได้อย่างมีนัยสำคัญ แต่เมื่อให้ในช่วงต้นพร้อมกับ LPS สารประกอบ 3 (พร้อมกับสารประกอบ 2) กลับทำให้ความเสียหายของหลอดเลือดรุนแรงขึ้น ซึ่งน่าจะเกิดจากการยับยั้ง eNOS ที่น่าสนใจคือ สารอนุพันธ์ที่มีขนาดใหญ่ขึ้นของสารประกอบ 3 หยุดการทำงานของ iNOS ด้วยกลไกที่แตกต่างจากสารประกอบ 3 เอง (ซึ่งเหนี่ยวนำการสูญเสียฮีม) แม้จะมีฤทธิ์อ่อนกว่าและยังไม่มีรายงานความจำเพาะก็ตาม
อะมิโนกัวนิดีน (สารประกอบ 4) เป็นสารทำให้หยุดทำงานแบบแข่งขันที่ขึ้นกับเวลาและอาศัยกลไกเฉพาะของ NOS ทุกไอโซฟอร์ม แต่อาจจับกับ iNOS ได้ดีกว่า คณะวิจัยของ Corbett รายงานว่าสารประกอบ 4 มีฤทธิ์เพิ่มความดันโลหิตแดงเฉลี่ยในหนูแรทน้อยกว่าสารประกอบ 2 ถึงประมาณ 40 เท่า ซึ่งบ่งชี้ถึงความจำเพาะบางส่วนต่อ iNOS เหนือกว่า eNOS ในสภาวะร่างกายจริง คณะวิจัยของ Misko ยืนยันความจำเพาะต่อ iNOS เหนือ nNOS ทั้งในระบบเซลล์และสารสกัดจากเซลล์ แต่ภายหลัง AstraZeneca รายงานว่าพบความจำเพาะเพียงเล็กน้อย ในสภาวะร่างกายจริง สารประกอบ 4 แสดงผลหลากหลายที่เกี่ยวข้องกับ iNOS รวมถึงความสามารถในการบรรเทาการเปลี่ยนแปลงเชิงลบของระบบหัวใจและหลอดเลือดในหนูแรทที่ได้รับเอนโดทอกซิน ช่วยเพิ่มอัตราการรอดชีวิตในแบบจำลองภาวะเอนโดทอกซีเมียของหนูเมาส์ และชะลอการเริ่มต้นและความรุนแรงของโรคในแบบจำลองโรคไขสันหลังอักเสบจากภูมิคุ้มกันตนเองของหนู (แบบจำลองของโรคปลอกประสาทเสื่อมแข็ง)
N6-(1-Iminoethyl)-L-lysine (L-NIL หรือสารประกอบ 5) เป็นสารทำให้หยุดทำงานแบบแข่งขันเช่นเดียวกับสารประกอบ 3 ซึ่งรายงานว่ามีฤทธิ์แรงกว่าในหลอดทดลอง (IC50 = 3.3 ไมโครโมลาร์สำหรับ iNOS ของหนูเมาส์) มากกว่าสารประกอบ 2, 3 และไนโตรอาร์จินีน โดยมีความจำเพาะต่อ iNOS เหนือ nNOS สูงถึง 28 เท่า ที่น่าพึงพอใจคือสารประกอบ 5 ช่วยลดความรุนแรงของภาวะข้อเสื่อมระยะแรกในสุนัขโดยไม่มีความเป็นพิษ ซึ่งอาจผ่านการลดการตายของเซลล์แบบอะพอพโทซิสในกระดูกอ่อน อย่างไรก็ตาม สารประกอบ 5 มีคุณสมบัติทางกายภาพที่ไม่พึงประสงค์ ซึ่งเป็นแรงผลักดันให้ Hallinan และคณะพัฒนาสารประกอบ 5 ในรูปโปรดรัก คือ เทตระโซลเอไมด์ L-NIL-TA (หรือ SC-51 สารประกอบ 6) แม้จะไม่มีฤทธิ์ในหลอดทดลองต่อ iNOS ของมนุษย์ แต่ยานี้สามารถดูดซึมทางปากได้ดีและถูกไฮโดรไลซ์เป็นสารประกอบ 5 ในร่างกาย โดยมีฤทธิ์ใกล้เคียงกับสารประกอบ 5 ในแบบจำลอง LPS เอนโดทอกซิน แบบจำลองการอักเสบจากคาราจีแนน และแบบจำลองข้ออักเสบที่เหนี่ยวนำด้วยสารเสริมฤทธิ์ในหนูแรทโดยไม่ทำให้ความดันโลหิตสูงขึ้น ที่น่าสนใจคือ สารประกอบ 6 ยังช่วยลดปริมาณ NO ที่หายใจออกทั้งในผู้ป่วยโรคหอบหืดและคนสุขภาพดี แต่ดูเหมือนจะยังไม่ได้รับการศึกษาทางคลินิกต่อสำหรับข้อบ่งชี้นี้ แม้ iNOS จะมีความเกี่ยวข้องกับพยาธิสภาพของโรคหอบหืดก็ตาม
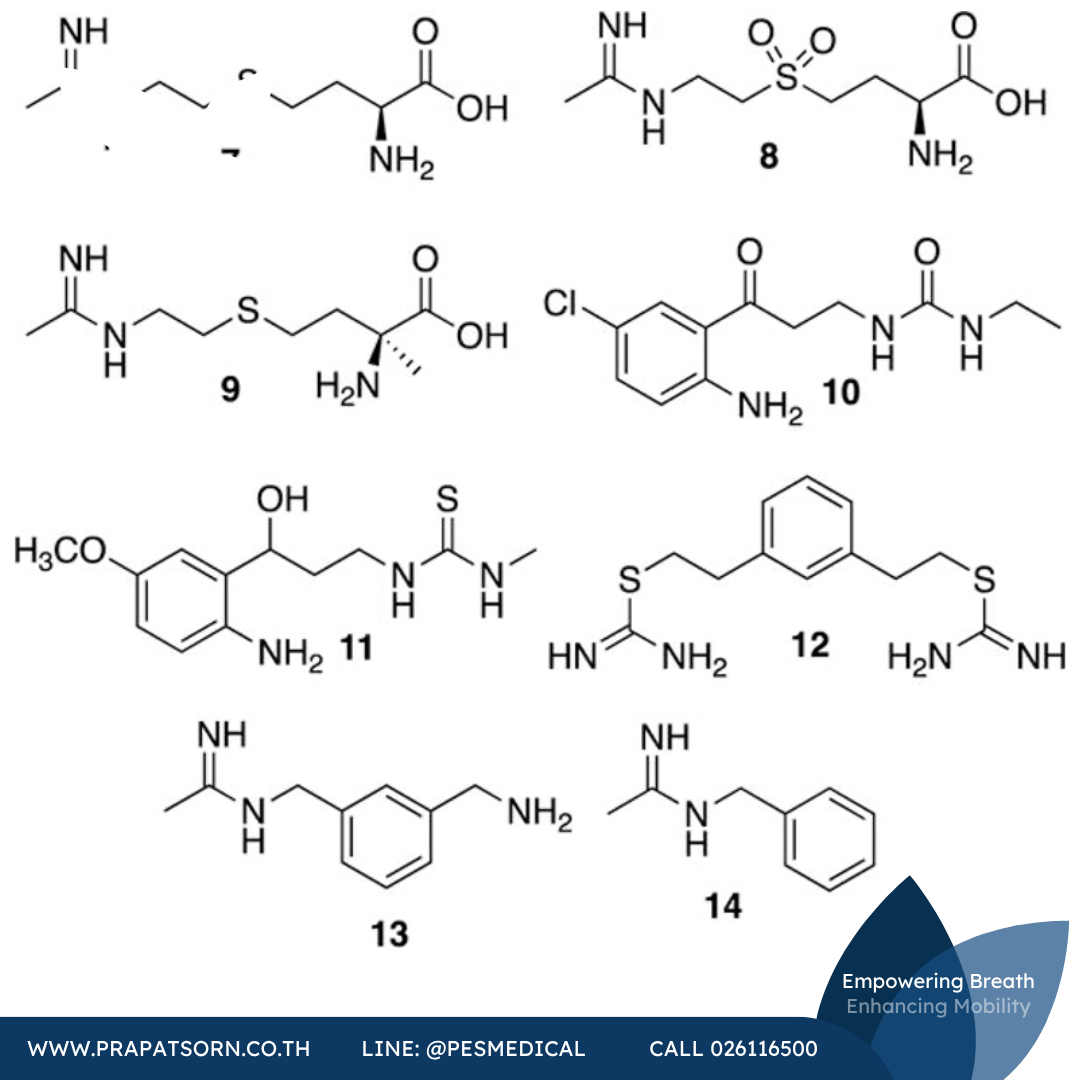
สารประกอบเอมิดีนของกรดอะมิโนอีกสองชนิดที่น่าสนใจคือสารประกอบ 7 และ 8 (พัฒนาโดยบริษัท Glaxo-Wellcome รู้จักกันในชื่อ GW274150 และ GW273629 ตามลำดับ) สารประกอบทั้งสองนี้มีฤทธิ์แรง (Kd < 90 นาโนโมลาร์, IC50 < 10 ไมโครโมลาร์) มีความจำเพาะสูง (i/e มากกว่า 80 เท่า, i/n มากกว่า 100 เท่า) และเป็นสารยับยั้ง (หรือสารทำให้หยุดทำงาน) ของ iNOS ทั้งในมนุษย์และสัตว์ฟันแทะที่ขึ้นกับเวลาและ NADPH สารประกอบเหล่านี้มีประสิทธิภาพในการยับยั้งการเพิ่มขึ้นของไนเตรต/ไนไตรต์ในพลาสมาในแบบจำลอง LPS ของหนูเมาส์ และยังไม่ยับยั้ง nNOS และ eNOS ในชิ้นเนื้อเยื่อหรือในร่างกายจริง สารประกอบ 7 แสดงฤทธิ์ระงับปวดในแบบจำลองความเจ็บปวดของหนูแรทหลายแบบ (การอักเสบของอุ้งเท้าที่เหนี่ยวนำด้วยสารเสริมฤทธิ์และการบาดเจ็บจากการกดทับเรื้อรัง) และลดการอักเสบของปอดที่เหนี่ยวนำด้วยคาราจีแนนในหนูแรทในลักษณะขึ้นกับขนาดยา (โดยเริ่มเห็นผลที่ 2.5 มก./กก.) สารประกอบ 7 ได้เข้าสู่การทดลองทางคลินิกระยะที่ 2 สำหรับโรคข้ออักเสบรูมาตอยด์และไมเกรน แม้จะแสดงความสามารถบางส่วนในการลดความหนาของเยื่อบุข้อและความหนาแน่นของหลอดเลือดในผู้ป่วยระยะ 2a แต่ก็ไม่ได้พัฒนาต่อไป นอกจากนี้ สารประกอบ 7 ยังไม่แสดงความสามารถในการป้องกันไมเกรนเมื่อให้เป็นยาป้องกันล่วงหน้า การพัฒนาทางคลินิกของสารประกอบ 7 ดูเหมือนจะยุติลงราวปี ค.ศ. 2012
สารอนุพันธ์ที่เกี่ยวข้องกัน คือสารยับยั้งแบบถาวรที่มีความจำเพาะสูงอย่างซินดูนิสแตต (SD6010 หรือสารประกอบ 9) ได้รับการศึกษาโดยบริษัทไฟเซอร์สำหรับการรักษาโรคข้อเสื่อม แม้จะแสดงประสิทธิภาพที่ดีในแบบจำลองสัตว์ทดลองของความเจ็บปวดจากการอักเสบ ความเจ็บปวดทางระบบประสาท และความเจ็บปวดที่สัมพันธ์กับข้อเสื่อม แต่สารประกอบ 9 กลับไม่สามารถลดความเจ็บปวดและการแคบลงของช่องว่างข้อในผู้ป่วยมนุษย์ได้ แม้ว่าจะไม่ได้วัดการเปลี่ยนแปลงอื่นๆ ของข้อ และยานี้มีความปลอดภัยและทนต่อยาได้ดี
สารประกอบยูเรีย ไทโอยูเรีย และไอโซไทโอยูเรียหลายชนิดก็ได้รับการศึกษาในฐานะสารยับยั้ง iNOS เช่นกัน คณะวิจัยของ Southan ได้ตีพิมพ์รายงานฉบับแรกเกี่ยวกับไอโซไทโอยูเรียอย่างง่ายที่ยับยั้ง iNOS ในปี ค.ศ. 1995 แต่โมเลกุลเหล่านี้ยังคงอยู่ระหว่างการศึกษาจนถึงปี ค.ศ. 2016 เมื่อไม่นานมานี้ ไคนูเรนามีนยูเรีย (เช่นสารประกอบ 10) และไทโอยูเรีย (สารประกอบ 11) ได้รับการพิสูจน์ว่าเป็นสารยับยั้งทั้ง iNOS หรือทั้ง iNOS และ nNOS ร่วมกัน โดยมีการยับยั้ง eNOS และความเป็นพิษต่อเซลล์ต่ำ แม้ฤทธิ์ของสารเหล่านี้จะยังไม่แรงมากนัก แต่การจำลองโมเลกุลสนับสนุนสมมติฐานว่าสารเหล่านี้แข่งขันกับแอล-อาร์จินีน สารประกอบบิสไอโซไทโอยูเรีย 12 (รู้จักกันในชื่อ PBITU) เป็นสารยับยั้ง iNOS ของมนุษย์ที่มีฤทธิ์แรงมาก (Ki = 47 นาโนโมลาร์) ด้วยความจำเพาะต่อ iNOS เหนือ eNOS สูงถึง 190 เท่า แต่แสดงความเป็นพิษเฉียบพลันสูงในสัตว์ฟันแทะ
การปรับปรุงโครงสร้างของสารประกอบ 12 นำไปสู่สารประกอบ 13 (1400W) ซึ่งเป็นสารยับยั้ง iNOS ที่มีฤทธิ์แรงมากและมีความจำเพาะสูง (ประมาณ 5,000 เท่าสำหรับ i/e และมากกว่า 250 เท่าสำหรับ i/n) ในเชิงกลไก สารประกอบ 13 แสดงฤทธิ์ยับยั้งแบบแข่งขันที่อ่อนต่อ eNOS และ nNOS เท่านั้น แต่การยับยั้ง iNOS พบว่าขึ้นกับเวลาและ NADPH ซึ่งบ่งชี้ถึงความเป็นไปได้ของการทำให้หยุดทำงาน (โดยมีค่า Kd ที่วัดได้ 7 นาโนโมลาร์) การศึกษาในภายหลังแสดงให้เห็นว่าสารประกอบ 13 ไม่ได้ถูกดัดแปลงโดยเอนไซม์หรือใช้เป็นสารตั้งต้น แต่ทำให้ iNOS หยุดทำงานผ่านกลไกที่สอดคล้องกับการสูญเสียฮีมในรูปบิลิเวอร์ดิน (เช่นเดียวกับสารประกอบ 3) การให้สารประกอบ 13 สามารถย้อนกลับความเสียหายของหลอดเลือดที่เหนี่ยวนำด้วย LPS ในหนูแรทได้ และสารประกอบ 13 ยังแสดงประสิทธิภาพในแบบจำลองความเจ็บปวดของสัตว์ทดลองด้วย การศึกษาล่าสุดชี้ให้เห็นว่าฤทธิ์ต้านการอักเสบของ 1400W อาจซับซ้อนกว่าเพียงแค่การยับยั้ง iNOS เนื่องจากในแบบจำลองความเจ็บปวดทางระบบประสาทของหนูแรท สัตว์ที่ได้รับการรักษากลับมีระดับไซโตไคน์ต้านความปวดเพิ่มขึ้น การศึกษา SAR ของสารเบนซามิดีนที่คล้ายคลึงกันโดยคณะวิจัยของ Amoroso เผยให้เห็นว่าสารประกอบ 14 ที่ถูกตัดโครงสร้างบางส่วนมีฤทธิ์แรงกว่าสารประกอบ 13 แต่มีความจำเพาะน้อยกว่า ซึ่งบ่งชี้ว่าหมู่อะมิโนเมทิลของสารประกอบ 13 ขัดขวางการจับกับ eNOS และจำเป็นต่อความจำเพาะที่สมบูรณ์
แม้บริษัท Merck, AstraZeneca, Searle/Pharmacia และกลุ่มวิจัยทางวิชาการหลายแห่งจะได้ศึกษาทั้งเอมิดีนของกรดอะมิโนและเอมิดีนอะโรมาติกในฐานะสารยับยั้ง iNOS แบบแข่งขัน แต่เอมิดีนแบบวงแหวน (cyclic amidines) กลับเป็นกลุ่มที่ได้รับการศึกษามากที่สุด ในบรรดา 2-iminoazaheterocycles อื่นๆ ที่คณะวิจัยของ Moore ศึกษา พบว่า 2-iminopiperidine (สารประกอบ 15) มีฤทธิ์แรงที่สุดต่อ iNOS ของมนุษย์ (IC50 = 1 ไมโครโมลาร์) โดยแสดงความสามารถในการยับยั้งการเพิ่มขึ้นของ NO ในพลาสมาของหนูแรทที่ได้รับ LPS เมื่อให้ทางปาก แต่ไม่ค่อยมีความจำเพาะต่อ nNOS หรือ eNOS มากนัก บริษัท Ono Pharmaceutical ค้นพบสาร iminopiperidine อีกชนิดหนึ่ง (ONO-1714 หรือสารประกอบ 16) ที่มีฤทธิ์ยับยั้ง iNOS แรงมาก (Ki = 1.88 นาโนโมลาร์) แต่มีความจำเพาะที่อ่อน (10 เท่า) ต่อ iNOS เหนือ eNOS สารประกอบนี้ยังถูกเปิดเผยในภายหลังว่าเป็นสารยับยั้ง nNOS ที่มีฤทธิ์แรงมากเช่นกัน สารประกอบ 16 ช่วยลดการบาดเจ็บของปอดในกระต่ายที่ได้รับเอนโดทอกซิน และลดภาวะกรดแลกติกในเลือดและความดันโลหิตต่ำในแบบจำลองภาวะช็อกจากเอนโดทอกซินในสุนัข แต่กลับทำให้เกิดความต้านทานของหลอดเลือดเพิ่มขึ้นที่ขนาดยาสูงในแบบจำลองสัตว์ดังกล่าว ซึ่งอาจเป็นผลจากการยับยั้ง eNOS 2-Iminopyrrolidines (เช่นสารประกอบ 17) ก็แสดงศักยภาพในฐานะสารยับยั้ง iNOS เช่นกัน (IC50 ของ iNOS มนุษย์ = 0.25 ไมโครโมลาร์) และมีฤทธิ์ในแบบจำลองภาวะเอนโดทอกซีเมียของสัตว์ฟันแทะ โดยมีความจำเพาะ i/e สูงกว่าสารประกอบ 15 และ 16 มาก แต่มีความจำเพาะ i/n ต่ำ (13 เท่า) การเติมหมู่ไฮดรอกซิล (เช่นในสารประกอบ 18) ทำให้ความจำเพาะ i/e เพิ่มขึ้น 3 เท่า และความจำเพาะ i/n เพิ่มขึ้น 2.5 เท่า
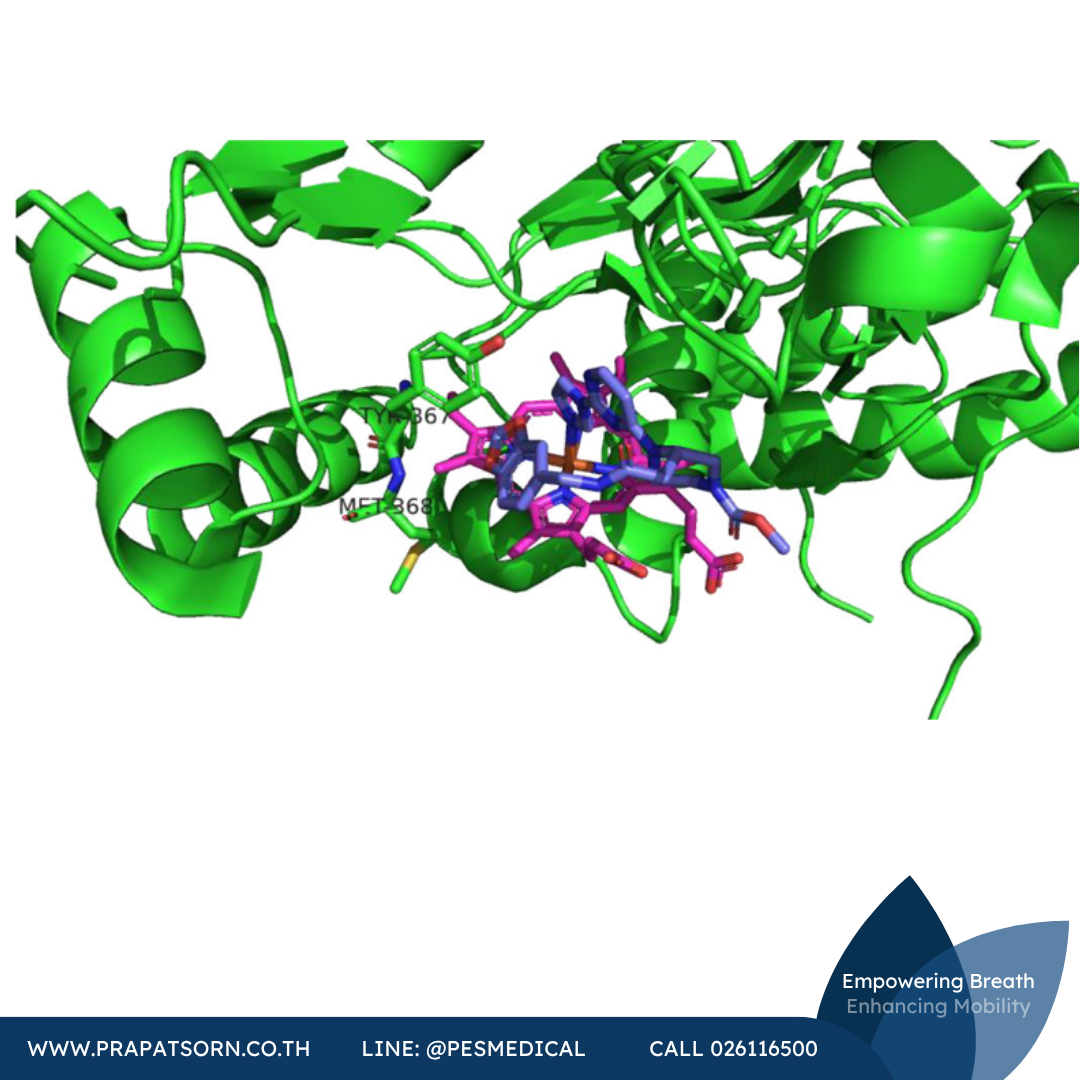
ในปี ค.ศ. 2001 บริษัท AstraZeneca รายงานว่าสาร 3,4-dihydro-1-isoquinolinamines (โดยเฉพาะ 3-phenyl-3,4-dihydro-1-isoquinolinamines) มีฤทธิ์ยับยั้ง iNOS สารประกอบ 19 มีค่า IC50 ต่อ iNOS ของมนุษย์ที่ 160 นาโนโมลาร์ ด้วยความจำเพาะ i/n ที่ 100 เท่า และความจำเพาะ i/e ที่ประมาณ 1,000 เท่า สารอนุพันธ์ของสารประกอบ 19 คือสไปโรไซคลิกเอไมด์ 20 (AR-C102222) ยังคงความจำเพาะ i/e สูงไว้ แต่มีความจำเพาะ i/n ลดลง อย่างไรก็ตาม สารประกอบนี้แสดงฤทธิ์ในร่างกายจริงสูงโดยไม่มีปัญหาระบบหัวใจและหลอดเลือด และมีผลปกป้องในแบบจำลองข้ออักเสบที่เหนี่ยวนำด้วยสารเสริมฤทธิ์ในหนูแรท สารประกอบ 20 ยังมีผลดีในแบบจำลองความเจ็บปวดและการอักเสบของหนูเมาส์หลายรูปแบบ (การอักเสบของหูและการบาดเจ็บของเส้นประสาทไขสันหลังและอุ้งเท้าหลัง)
แม้โครงสร้างผลึกเอกซเรย์ของ iNOS จะมีให้ศึกษามานานกว่า 20 ปีแล้ว แต่การออกแบบสารยับยั้ง iNOS ส่วนใหญ่ก็ยังไม่ได้อาศัยโครงสร้างเป็นหลักอย่างชัดเจน ข้อยกเว้นที่น่าสนใจคือวิธี anchored plasticity ของ AstraZeneca ในวิธีนี้ มีการผสมผสานระหว่างผลึกศาสตร์เอกซเรย์และการศึกษาการกลายพันธุ์เพื่อค้นหาช่องจำเพาะของ iNOS ที่เปิดออกเมื่อสารยับยั้งจับตัว อันเป็นผลจากการเปลี่ยนแปลงเชิงโครงสร้างของเอนไซม์ พื้นที่นี้จึงถูกนำมาใช้ออกแบบสารประกอบใหม่ โดยใช้กลยุทธ์ที่หมู่ฟังก์ชันซึ่งทราบว่ามีฤทธิ์ยับยั้ง iNOS (เช่นเอมิดีน) ถูก “ยึดตรึง” ไว้ในตำแหน่งจับ แล้วขยายส่วนอื่นออกไปเพื่อเข้าถึงช่องจำเพาะที่เกิดจากการปรับตัวรับนี้ ด้วยวิธีนี้ สารประกอบ 21 ซึ่งมี IC50 400 นาโนโมลาร์ (ต่อ iNOS มนุษย์) และความจำเพาะ n/e 200 เท่า จึงถูกออกแบบขึ้น ในโครงสร้างผลึกเอกซเรย์ของสารประกอบ 21 ที่จับกับ iNOS ของหนูเมาส์ พบว่าหมู่ไทโอฟีนคาร์บอกซาไมดีนของสารประกอบ 21 เลียนแบบแอล-อาร์จินีนและสร้างสะพานเกลือกับกรดกลูตามิกที่ตำแหน่งเร่งปฏิกิริยา
ภาพที่ 8: โครงสร้างผลึกเอกซเรย์ของสารประกอบ 21 (สีม่วง) ที่จับกับตำแหน่งเร่งปฏิกิริยาของ iNOS หนูเมาส์
2-Aminopyridine เป็นสารเลียนแบบแอล-อาร์จินีนอีกกลุ่มหนึ่งที่ได้รับการศึกษาเมื่อไม่นานมานี้สำหรับทั้ง iNOS และ nNOS AstraZeneca รายงานว่าอะมิโนไพริดีนอย่างง่ายสามารถต้านฤทธิ์แอล-อาร์จินีนใน iNOS ได้ แม้สารประกอบส่วนใหญ่ในกลุ่มนี้จะไม่มีความจำเพาะ มีความจำเพาะอ่อน หรือมีฤทธิ์ต่ำกว่าเอมิดีนแบบวงแหวนอิ่มตัว (เช่นสารประกอบ 15) ทั้งในหลอดทดลองและในร่างกายจริง 4-Methylaminopyridine (สารประกอบ 22 หรือ 4-MAP) เป็นสารยับยั้ง iNOS ที่ไม่มีความจำเพาะ แต่แสดงฤทธิ์ในแบบจำลองภาวะเอนโดทอกซีเมียและความเจ็บปวดของสัตว์ฟันแทะ แม้ขนาดยาที่จำเป็นสำหรับฤทธิ์ระงับปวดจะต่ำกว่าขนาดที่จำเป็นสำหรับประสิทธิภาพในแบบจำลองภาวะช็อกจากเอนโดทอกซิน ซึ่งบ่งชี้ว่าคุณสมบัติระงับปวดบางส่วนน่าจะเป็นผลจากการยับยั้ง nNOS
อะมิโนไพริดีนที่มีการแทนที่สองตำแหน่ง (สารประกอบ 23) มีฤทธิ์แรงกว่าสารประกอบ 22 ประมาณ 6 เท่า และมีความจำเพาะที่ดีขึ้น แต่โดยทั่วไปแล้วอะมิโนไพริดีนที่มีโครงสร้างซับซ้อนกว่าจะมีประสิทธิภาพดีกว่า จากการปรับปรุงโครงสร้าง จึงค้นพบสารประกอบ AR-C133057XX (สารประกอบ 24) ซึ่งเป็นสารอนุพันธ์พิเพอริดีนเอไมด์ที่มีฤทธิ์แรงทั้งในร่างกายจริงและในหลอดทดลอง (IC50 = 71 นาโนโมลาร์ต่อ iNOS มนุษย์) และมีความจำเพาะสูงมากต่อ iNOS เหนือ eNOS (ยับยั้ง eNOS น้อยกว่าร้อยละ 20 ที่ความเข้มข้น 100 ไมโครโมลาร์) โครงสร้างผลึกเอกซเรย์เผยให้เห็นว่าหมู่อะมิโนไพริดีนของสารประกอบ 24 พลิกกลับ 180 องศาเมื่อเทียบกับสารประกอบ 22 ซึ่งผู้วิจัยเสนอว่าทำให้หมู่พิเพอริดีนและเบนโซไนไตรล์สามารถจับกับบริเวณจำเพาะเพิ่มเติมของ iNOS ได้
สารอนุพันธ์ไพริดีนที่ไม่ธรรมดาชนิดหนึ่งคืออิมิดาโซไพริดีน 25 (BYK191023) ซึ่งถูกค้นพบในปี ค.ศ. 2005 จากการคัดกรองแบบ high-throughput สองปีต่อมา มีการพิสูจน์ว่าเป็นสารยับยั้งแบบแข่งขันที่ขึ้นกับเวลาและไม่ผันกลับในสภาวะที่มี NADPH ซึ่งน่าจะก่อให้เกิดการสูญเสียฮีม สารประกอบ 25 มีความจำเพาะสูงต่อ iNOS มาก (ทั้งในการทดสอบเอนไซม์ เซลล์ และระบบอวัยวะแยก) และยังได้รับการทดสอบในแบบจำลองสัตว์ทดลองหลายรูปแบบ นอกจากการลดไนไตรต์/ไนเตรตในพลาสมาและการรักษาความดันโลหิตในหนูแรทที่ได้รับ LPS แล้ว สารประกอบ 25 ยังช่วยเพิ่มการไหลเวียนเลือดของอวัยวะและลดความดันในหลอดเลือดปอดสูงและภาวะกรดในแกะที่มีภาวะติดเชื้อในกระแสเลือด (เมื่อเทียบกับแกะที่ได้รับนอร์อิพิเนฟริน) แต่ยานี้ไม่ได้เพิ่มอัตราการรอดชีวิตโดยรวม
สารยับยั้งการรวมตัวเป็นไดเมอร์ (Dimerization Inhibitors)
กลไกที่สองซึ่งการพัฒนาสารยับยั้งได้มุ่งเน้นคือการยับยั้งการรวมตัวเป็นไดเมอร์ที่มีฤทธิ์ของ iNOS เนื่องจากบริเวณรอยต่อของไดเมอร์อยู่ที่ปลายสุดของตำแหน่งเร่งปฏิกิริยา สารยับยั้งที่ยึดเกาะในตำแหน่งเร่งปฏิกิริยาและขัดขวางการรวมตัวเป็นไดเมอร์จึงเป็นที่ต้องการ เป็นที่ทราบกันมานานแล้วว่าสารกลุ่มอิมิดาโซล (imidazoles) สามารถประสานพันธะกับเหล็กในฮีมของ NOS ทุกไอโซฟอร์มได้ ในขณะที่อิมิดาโซลขนาดเล็กจะช่วยเสริมความเสถียรของการรวมตัวเป็นไดเมอร์ของ iNOS แต่อิมิดาโซลที่มีขนาดใหญ่ (เช่น สารต้านเชื้อราโคลไตรมาโซล หรือสารประกอบ 26) กลับป้องกันการประกอบตัวเป็นไดเมอร์
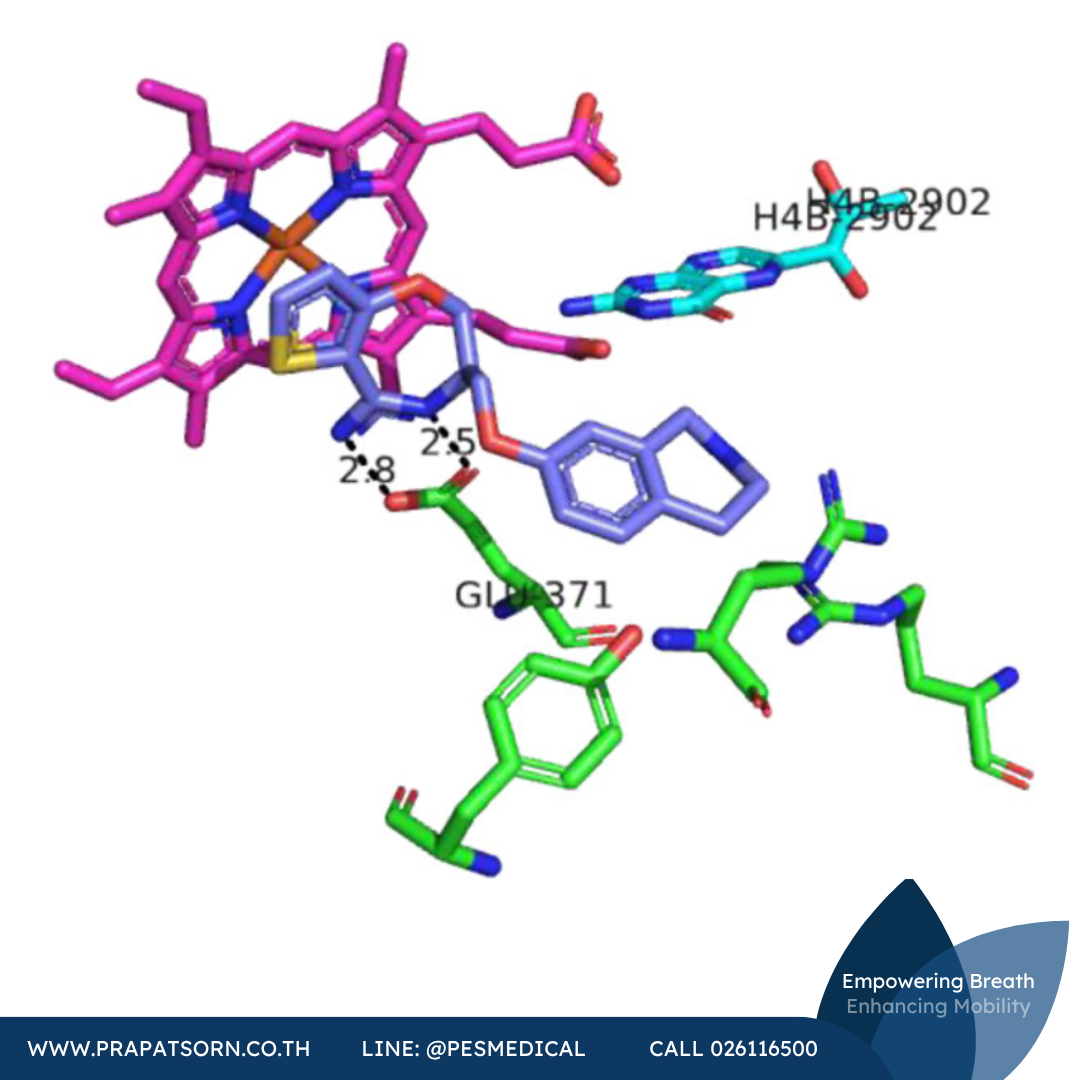
หลังจากการค้นพบนี้ Berlex Bioscience ได้ค้นพบสารกลุ่มไพริมิดินิลอิมิดาโซล (เช่นสารประกอบ 27) ผ่านเคมีเชิงผสมผสาน สารประกอบเหล่านี้จับกับ iNOS มอนอเมอร์ของมนุษย์ด้วยความสัมพันธ์สูง (Ki = 2.2 นาโนโมลาร์สำหรับสารประกอบ 27) และแสดงฤทธิ์ในแบบจำลอง LPS ของสัตว์ฟันแทะ โครงสร้างผลึกเอกซเรย์ของสารประกอบ 27 ที่จับกับ iNOS มอนอเมอร์ของหนูเมาส์ (ที่ตัดกรดอะมิโน 114 ตัวแรกจากปลาย N ออก) บ่งชี้ว่าสารประกอบเหล่านี้ออกฤทธิ์โดยการประสานพันธะกับเหล็ก ปรับโครงแบบเป็นรูปตัวยู และแทนที่ตำแหน่ง Glu371 ออกจากตำแหน่งเร่งปฏิกิริยา การรบกวนหรือทำให้เฮลิกซ์ของกรดอะมิโนตำแหน่งนี้ (เฮลิกซ์ 7a) ไม่เป็นระเบียบ จะรบกวนบริเวณรอยต่อของการรวมตัวเป็นไดเมอร์ตามมา การศึกษาเชิงกลไก (สเปกโทรสโกปีและการแข่งขันจับตัว) โดยคณะวิจัยของ Blasko ยืนยันในภายหลังว่าไพริมิดินิลอิมิดาโซลออกฤทธิ์โดยการประสานพันธะกับศูนย์กลางฮีมของ iNOS มอนอเมอร์และยับยั้งการรวมตัวเป็นไดเมอร์แบบอัลโลสเตอริก โดยเสนอว่าความจำเพาะที่สังเกตได้สะท้อนถึงจลนศาสตร์การประกอบตัวเป็นไดเมอร์ที่แตกต่างกันใน eNOS และ nNOS (เทียบกับ iNOS) เมื่อไม่นานมานี้ยังมีการแสดงให้เห็นว่าไพริมิดินิลอิมิดาโซลสามารถแยกไดเมอร์ที่มีฤทธิ์ออกเป็นมอนอเมอร์ในเซลล์ได้ และสารประกอบเชิงซ้อนระหว่างมอนอเมอร์กับสารยับยั้งนี้ไม่สามารถผันกลับได้ และไม่สามารถเปลี่ยนกลับเป็นไดเมอร์ที่มีฤทธิ์ได้แม้จะเติมแอล-อาร์จินีนหรือ H4B เข้าไป
ภาพที่ 11: โครงสร้างผลึกเอกซเรย์ของสารประกอบ 27 (สีม่วง) ที่จับกับ iNOS มอนอเมอร์ แสดงการประสานพันธะกับฮีม (สีชมพู) และการรบกวนเฮลิกซ์ที่มี Glu371
จากการศึกษาผลึกเอกซเรย์ สารประกอบโฮโมโพรลีน 28 ก็ป้องกันการรวมตัวเป็นไดเมอร์เช่นกันโดยการแทนที่เฮลิกซ์ 7a ของ iNOS สารประกอบนี้สามารถดูดซึมทางปากได้และเป็นสารยับยั้งที่ดีเยี่ยมในเซลล์ (IC50 = 290 พิโคโมลาร์) ซึ่งน่าจะออกฤทธิ์โดยจับกับ iNOS มอนอเมอร์ที่เพิ่งก่อตัวขึ้นหรือหลังจากก่อตัวขึ้นไม่นาน สารประกอบ 28 ยังลดคะแนนความรุนแรงของข้ออักเสบทางคลินิกในหนูแรทที่มีข้ออักเสบซึ่งเหนี่ยวนำด้วยสารเสริมฤทธิ์ด้วย
นอกเหนือจากงานของ Berlex Biosciences แล้ว บริษัท Kalypsys Inc. ก็มีบทบาทสำคัญในการพัฒนาสารยับยั้งการรวมตัวเป็นไดเมอร์ของ iNOS เช่นกัน ในปี ค.ศ. 2009 มีการคัดกรองแบบเซลล์ที่มีความเร็วสูงมาก (ultra high-throughput) กับสารประกอบที่ไม่ใช่แอล-อาร์จินีนและไม่ใช่อิมิดาโซล (เนื่องจากกังวลเรื่องการยับยั้งไซโตโครม) และพบสารตั้งต้นควิโนลิโนน 29 จากนั้นจึงมีการปรับปรุงโครงสร้างเพื่อเพิ่มฤทธิ์ต่อ iNOS ของหนูเมาส์ (สำหรับการใช้ในสัตว์ทดลอง) และพบสารตัวนำใหม่ 30 (KLYP596) และ 31 ที่มีฤทธิ์และความจำเพาะดีขึ้น โดยสารประกอบ 30 มีค่า EC50 ที่ 11 นาโนโมลาร์ และมีความจำเพาะสูงกว่าในสองตัว (i/e เท่ากับ 2,300 และ i/n เท่ากับ 210) ในการทดสอบแบบเซลล์ การตรวจด้วยเจลอิเล็กโทรโฟรีซิสและอิมมูโนบล็อตของเอนไซม์ที่แยกได้จากเซลล์ที่ผ่านการรักษาเผยให้เห็นการสูญเสียของ iNOS แบบไดเมอร์ที่ขึ้นกับขนาดยา การศึกษาทางชีวเคมีเผยให้เห็นว่าการยับยั้งนี้ไม่ไวต่อการเติมแอล-อาร์จินีนและ H4B แต่การจับตัวของสารประกอบ 30 ก็ไม่ก่อให้เกิดการเปลี่ยนแปลงทางสเปกตรัมที่สอดคล้องกับการประสานพันธะฮีมแบบอิมิดาโซล แม้สารประกอบ 30 และ 31 จะมีชีวประสิทธิผลทางปากที่ดี แต่ก็มีอัตราการกำจัดยาสูงและครึ่งชีวิตสั้นในร่างกายจริง ถึงกระนั้น สารประกอบทั้งสองก็ช่วยลดระดับไนเตรตในพลาสมาของหนูเมาส์ที่ได้รับ LPS และลดพฤติกรรมแสดงความเจ็บปวดในแบบจำลองฟอร์มาลินและการบาดเจ็บจากการกดทับเส้นประสาทเรื้อรังของหนูเมาส์
เพื่อปรับปรุงเภสัชจลนศาสตร์ นักวิจัยของ Kalypsys ยังได้ศึกษาควิโนลิโนน-เบนซิมิดาโซลและอิมิดาโซไพราซีนในฐานะสารอนุพันธ์ที่จำกัดโครงแบบของสารประกอบ 29 และหลังจากการวิเคราะห์ SAR ก็ค้นพบสารอนุพันธ์ไซโคลบิวทิล 32 (KD7332) ซึ่งเป็นสารยับยั้งคู่ iNOS/nNOS ที่มีฤทธิ์แรง สารประกอบ 32 มีอัตราการกำจัดยาลดลงในร่างกายจริง และแสดงฤทธิ์ต้านความเจ็บปวดในแบบจำลองฟอร์มาลินของหนูเมาส์และแบบจำลอง Chung ของความเจ็บปวดทางระบบประสาท โดยมีดัชนีการรักษา (therapeutic index) ที่กว้าง สารประกอบอีกชนิดหนึ่งของ Kalypsys ที่มีฤทธิ์แรง (IC50 = 3 นาโนโมลาร์) และมีความจำเพาะสูง (nNOS IC50 = 2.4 ไมโครโมลาร์, eNOS = มากกว่า 30 ไมโครโมลาร์) คือสารประกอบ 33 (KD7040) ซึ่งถูกอธิบายว่าเป็นสารยับยั้งการรวมตัวเป็นไดเมอร์ของ iNOS ที่ซึมผ่านผิวหนังได้และมีความจำเพาะสูง สารนี้ได้รับการศึกษา (ระยะที่ 1) สำหรับการใช้ทาเฉพาะที่ในการรักษาอาการปวดเส้นประสาทภายหลังงูสวัด (post-herpetic neuralgia) แต่การพัฒนาดูเหมือนจะยุติลงราวปี ค.ศ. 2008
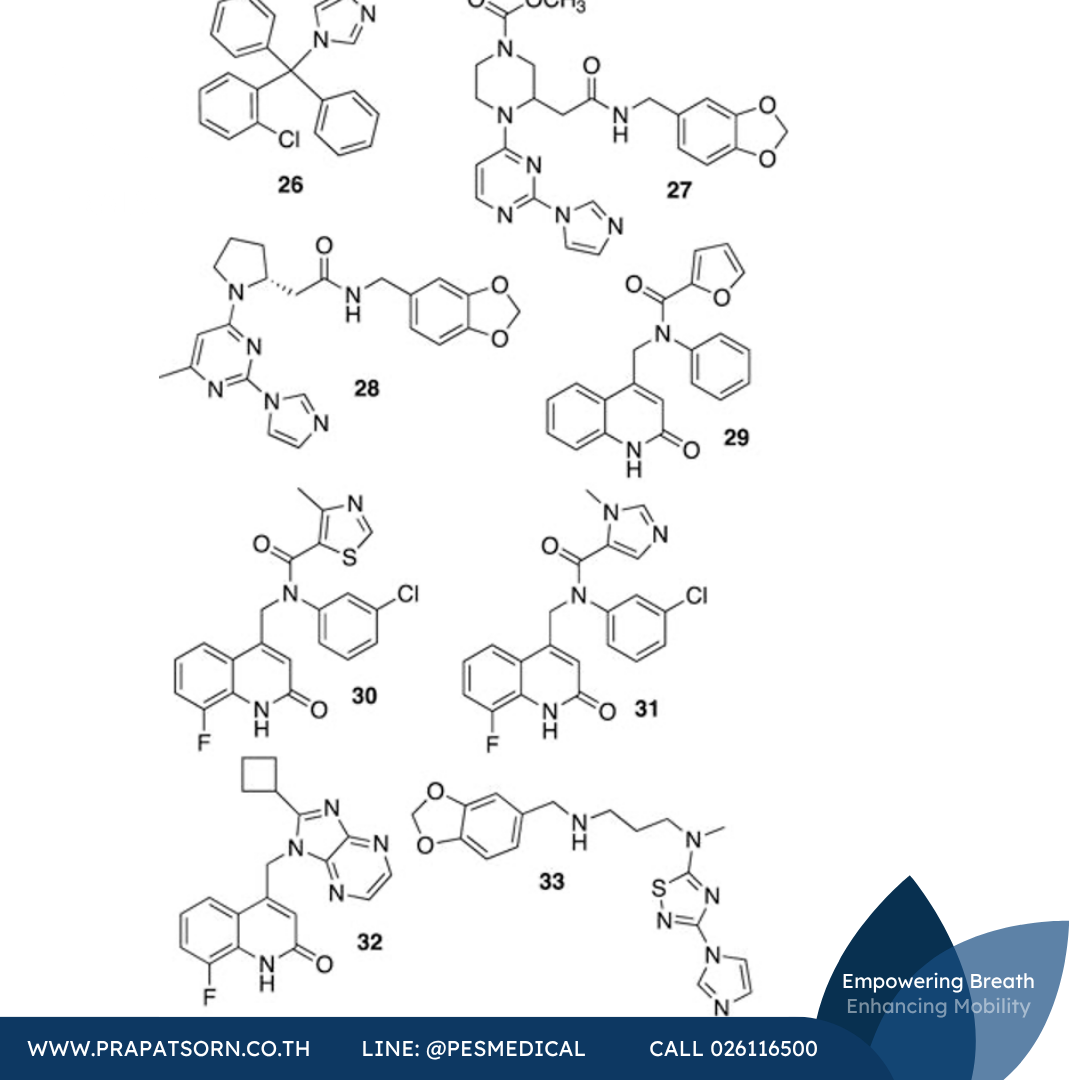
ภาพที่ 10: โครงสร้างทางเคมีของสารยับยั้งการรวมตัวเป็นไดเมอร์ของ iNOS ชนิดต่างๆ (สารประกอบ 26–33)
ความล้มเหลวทางคลินิก
ข้อบ่งชี้แบบดั้งเดิมสำหรับการยับยั้ง iNOS คือภาวะติดเชื้อในกระแสเลือด (การอักเสบทั่วร่างกาย) และความเจ็บปวด อย่างไรก็ตาม ความพยายามในการรักษาภาวะติดเชื้อในกระแสเลือดและภาวะช็อกจากการติดเชื้อด้วยสารยับยั้ง iNOS ในสภาวะทางคลินิกกลับประสบความสำเร็จอย่างจำกัด แม้จะมีการแสดงผลดีบางประการ แต่การยับยั้ง iNOS แบบไม่มีความจำเพาะ (เช่นด้วยสารประกอบ 2) กลับสามารถเพิ่มความผิดปกติของหัวใจและอัตราการเสียชีวิตในผู้ป่วยภาวะช็อกจากการติดเชื้อได้ มีข้อเสนอว่า eNOS อาจมีบทบาทปกป้องในภาวะติดเชื้อในกระแสเลือด (ดังที่แสดงให้เห็นในการศึกษาการแสดงออกมากเกินไปของ eNOS ในหนูเมาส์) ดังนั้นการยับยั้งแบบไม่มีความจำเพาะจึงอาจก่อให้เกิดอันตรายได้ การยับยั้ง iNOS แบบจำเพาะก่อให้เกิดผลข้างเคียงน้อยกว่า แต่ก็ยังคงเป็นที่ถกเถียงสำหรับข้อบ่งชี้นี้ นอกจากนี้ หนูที่ถูกน็อกเอาต์ยีน iNOS (หรือหนูที่ได้รับสารประกอบ 1, 2 หรือ 3) ก็ไม่ได้รับการปกป้องจากการเกิดลักษณะเอนโดทอกซีเมีย ผลนี้ยังพบในเซลล์ตับด้วย Bredan และ Cauwels ยังเสนอว่าสารที่จับ NO หรือควบคุมผลกระทบปลายน้ำของ NO อาจมีประสิทธิภาพมากกว่าการยับยั้ง iNOS โดยตรง
แม้จะมีประสิทธิภาพในแบบจำลองสัตว์ทดลอง แต่ก็ยังไม่มีสารยับยั้ง iNOS ตัวใดได้รับการรับรองสำหรับการรักษาความเจ็บปวด การทดลองด้านโรคข้ออักเสบล้มเหลวซ้ำแล้วซ้ำเล่า และฤทธิ์ต้านความเจ็บปวดในแบบจำลองสัตว์ทดลองก็ไม่สามารถแปลผลไปสู่มนุษย์ในการทดลองทางคลินิกได้ ประการแรก เป็นไปได้ว่าบทบาทของ NO ในความเจ็บปวดอาจซับซ้อนกว่าแนวคิดง่ายๆ ที่ว่า “NO มากเกินไปย่อมไม่ดี” (ดังที่เสนอไว้สำหรับกรณีภาวะติดเชื้อในกระแสเลือด) ประการที่สอง เป็นไปได้ว่าการยับยั้ง iNOS อาจมีประโยชน์จริง แต่การพัฒนายารักษาความเจ็บปวด (ซึ่งไม่ได้จำกัดเฉพาะ iNOS) เต็มไปด้วยความล้มเหลว ส่วนหนึ่งเป็นเพราะข้อจำกัดของวิธีทดสอบ และอีกส่วนหนึ่งเป็นเพราะความเจ็บปวดเป็นเรื่องอัตวิสัยที่ทั้งส่งผลกระทบต่อและได้รับผลกระทบจากพฤติกรรมที่ซับซ้อนอื่นๆ ทั้งในสัตว์ทดลองและมนุษย์
อีกพื้นที่หนึ่งที่การยับยั้ง iNOS ดูเหมือนจะล้มเหลวคือการรักษาภาวะช็อกจากหัวใจที่สัมพันธ์กับกล้ามเนื้อหัวใจตาย การศึกษาในระยะแรกชี้ให้เห็นว่า iNOS มีบทบาทสำคัญในภาวะหัวใจล้มเหลว เนื่องจากการแสดงออกมากเกินไปของ iNOS ในเซลล์หัวใจของหนูเมาส์ส่งผลให้เซลล์หัวใจตาย อย่างไรก็ตาม ในการทดลองทางคลินิกที่รู้จักกันดีที่สุดสำหรับภาวะช็อกจากหัวใจในผู้ป่วยกล้ามเนื้อหัวใจตาย (การทดลอง TRIUMPH) สารประกอบ 2 ไม่สามารถลดความรุนแรงของภาวะหัวใจล้มเหลวหรืออัตราการเสียชีวิตได้ และยังส่งผลให้เกิดความดันโลหิตสูงอีกด้วย มีการตั้งสมมติฐานว่าการขาดความจำเพาะหรือขนาดยาที่ต่ำเกินไปอาจเป็นสาเหตุของความล้มเหลวเหล่านี้ การศึกษาล่าสุด (ปี ค.ศ. 2015) ในสุกรที่ได้รับการช่วยชีวิต พบว่าทั้งการยับยั้ง NOS แบบรวม (โดยใช้สารประกอบ 1) และการยับยั้ง iNOS แบบจำเพาะมากขึ้น (โดยใช้สารประกอบ 4) ไม่ได้ช่วยเพิ่มอัตราการรอดชีวิตหรือความผิดปกติของกล้ามเนื้อหัวใจภายหลังภาวะหัวใจเต้นผิดจังหวะแบบสั่นพลิ้วในภาวะหัวใจหยุดเต้น (ซึ่งเป็นช่วงที่ระดับ iNOS เพิ่มขึ้น) นอกจากนี้ การสังเกตว่าการยับยั้งแบบไม่มีความจำเพาะส่งผลเสียต่อความดันโลหิตและการทำงานของหัวใจในการศึกษานี้ ยังชี้ให้เห็นว่า NO อาจมีความสำคัญอย่างแท้จริงต่อการฟื้นตัวที่เหมาะสมภายหลังเหตุการณ์เกี่ยวกับหัวใจ
ทิศทางใหม่สำหรับการรักษาด้วยการยับยั้ง iNOS
แม้ความสนใจส่วนใหญ่จะมุ่งเน้นที่ภาวะช็อกจากการติดเชื้อในกระแสเลือดและความเจ็บปวดประเภทต่างๆ แต่ก็เป็นไปได้ว่าโรคอื่นๆ ที่มีองค์ประกอบของการอักเสบในระดับมากก็อาจได้รับประโยชน์จากสารยับยั้ง iNOS เช่นกัน อันที่จริง มีการศึกษาจำนวนมากที่บ่งชี้ว่าการยับยั้ง iNOS อาจมีประโยชน์ทางการรักษาในวงกว้างขึ้นสำหรับโรคอื่นๆ รวมถึงโรคทางระบบประสาท เบาหวาน มะเร็ง และโรคระบบทางเดินหายใจ ซึ่งจะสรุปโดยย่อดังต่อไปนี้
แม้ไอโซฟอร์ม NOS ที่ได้รับการศึกษามากที่สุดในฐานะเป้าหมายของสารต้านความเสื่อมของระบบประสาทจะเป็น nNOS แต่หลักฐานก็ชี้ให้เห็นว่าการยับยั้ง iNOS ก็อาจเป็นประโยชน์ต่อการรักษาหรือป้องกันความผิดปกติทางระบบประสาทเสื่อม เช่น โรคพาร์กินสัน หรือความเสียหายของเซลล์ประสาทที่สัมพันธ์กับโรคหลอดเลือดสมองด้วย ตัวอย่างเช่น การปิดกั้นการแสดงออกของ iNOS (ด้วย siRNA) ช่วยลดอาการแบบพาร์กินสันในสัตว์ทดลองที่ได้รับสารพิษต่อระบบประสาท 6-OHDA สารประกอบ 4 แสดงผลดีในแบบจำลองสัตว์ทดลองของโรคหลอดเลือดสมอง เช่น ความสามารถในการลดปริมาตรของเนื้อสมองที่ตายในลูกหนูแรทที่มีภาวะขาดออกซิเจนและขาดเลือด และในหนูแรทตัวเต็มวัย แม้การยับยั้ง iNOS จะเชื่อว่ามีส่วนรับผิดชอบเพียงบางส่วนของผลที่สังเกตได้ในการศึกษาหลัง กลไกอีกประการหนึ่งที่อาจเป็นไปได้คือการยับยั้งเอนไซม์พอลิเอมีนออกซิเดสโดยสารประกอบ 4 การยับยั้ง iNOS แบบจำเพาะ (เช่นด้วยสารประกอบ 7) ยังพบว่ามีฤทธิ์ปกป้องเซลล์ประสาท ป้องกันการอักเสบของระบบประสาทในแบบจำลองพาร์กินสันแบบ 6-OHDA ในสัตว์ฟันแทะที่ขนาดยา 10 มก./กก. แม้ขนาดยาที่สูงขึ้นจะดูเหมือนมีผลน้อยลงก็ตาม
สารฟลาโวนอลจากธรรมชาติอย่างเควอซิทิน (สารประกอบ 34) ป้องกันทั้งการตายของเซลล์แบบอะพอพโทซิสที่เหนี่ยวนำด้วย 6-OHDA ในเซลล์เพาะเลี้ยงคล้ายเซลล์ประสาท PC12 และการตายของเซลล์ประสาทในปลาม้าลาย ปรากฏการณ์นี้พบเฉพาะในตัวอ่อนปลาม้าลายระยะแรกเท่านั้น ซึ่งบ่งชี้ว่าสารประกอบนี้อาจไม่สามารถผ่านกำแพงกั้นเลือด-สมองได้อย่างมีประสิทธิภาพ เควอซิทินยังลดการแสดงออกของ iNOS ในเซลล์เพาะเลี้ยง PC12 ด้วย (และมีข้อเสนอว่าฤทธิ์ปกป้องเซลล์ประสาทบางส่วนอาจมาจากการลดการแสดงออกของ iNOS) แต่สารประกอบ 34 ก็สามารถลดการแสดงออกของไซโตไคน์ก่อการอักเสบชนิดอื่นได้ด้วย ดังนั้นผลของมันจึงอาจมีหลายแง่มุม
รายงานในปี ค.ศ. 2014 ฉบับหนึ่งกล่าวถึงสารประกอบกลุ่มคล้ายผลิตภัณฑ์ธรรมชาติที่ค้นพบผ่านการคัดกรองเสมือนจริงแบบ high-throughput สารประกอบชนิดหนึ่ง (บิส-อินโดล 35) ถูกเสนอว่าเป็นสารยับยั้งแบบแข่งขันของการผลิต NO ในแมคโครฟาจที่ถูกกระตุ้น สารประกอบ 35 ยังแสดงฤทธิ์ปกป้องเซลล์ประสาทบางส่วนในตัวอ่อนปลาม้าลายที่ได้รับสารพิษต่อระบบประสาท MPTP แม้ผลเหล่านี้จะยังไม่มีความสัมพันธ์ที่ชัดเจนกับการยับยั้ง iNOS โดยเฉพาะก็ตาม
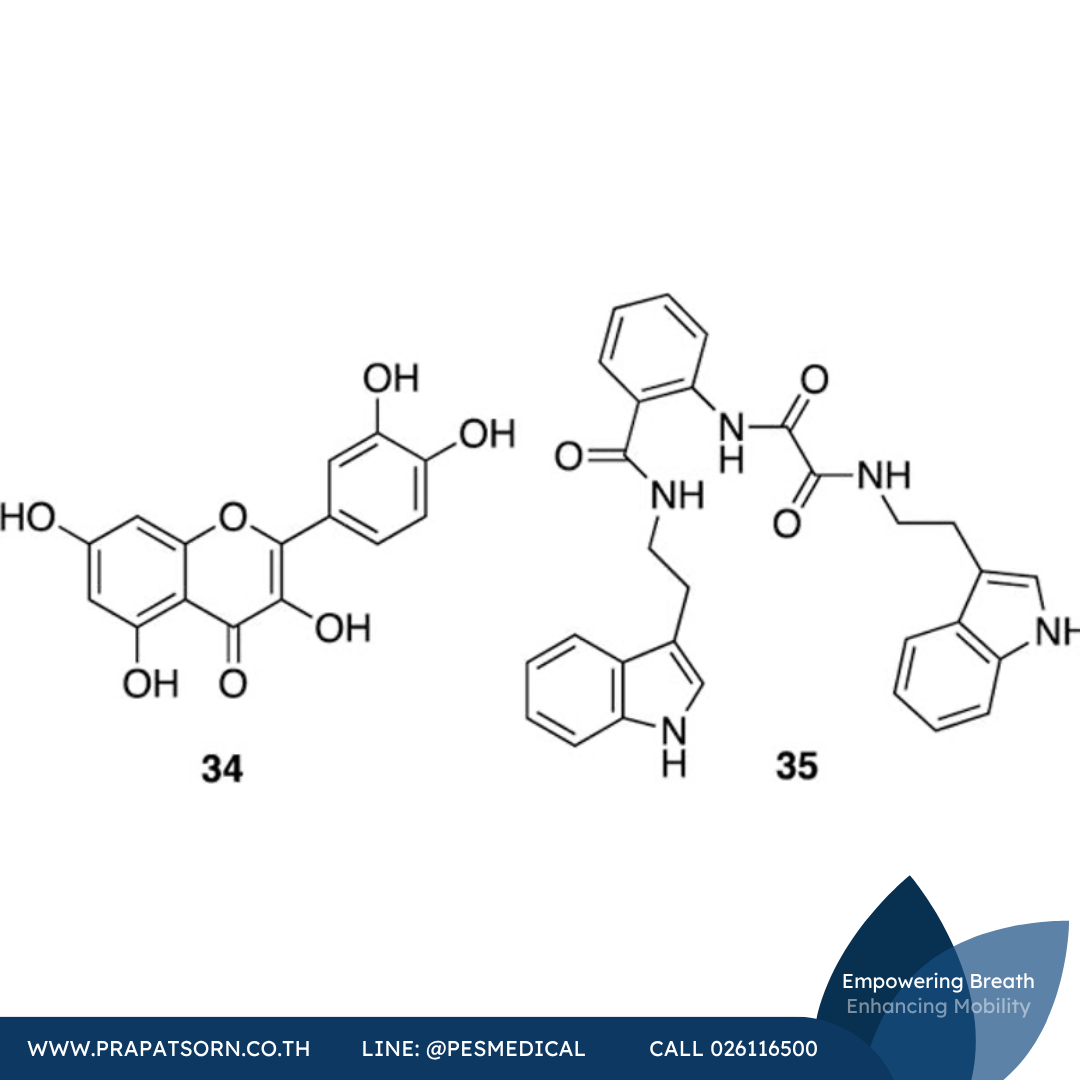
ภาพที่ 12: โครงสร้างทางเคมีของสารยับยั้ง iNOS ที่มีศักยภาพปกป้องเซลล์ประสาท ได้แก่ เควอซิทิน (สารประกอบ 34) และบิส-อินโดล (สารประกอบ 35)
การยับยั้ง iNOS ยังได้รับการศึกษาเมื่อไม่นานมานี้ในฐานะเป้าหมายที่น่าสนใจสำหรับโรคลมชัก Uzüm และคณะแสดงให้เห็นในหนูแรทว่าโรคลมชัก (แสดงออกเป็นอาการชักแบบเกร็งกระตุกในระยะยาว) เช่นเดียวกับภาวะติดเชื้อในกระแสเลือด มีความสัมพันธ์กับการอักเสบทั่วร่างกาย (รวมถึงระดับ iNOS ที่เพิ่มขึ้นในตับและไต) การศึกษาอื่นๆ แสดงให้เห็นว่าการยับยั้ง iNOS อาจเป็นประโยชน์ในโรคลมชัก ตัวอย่างเช่น อะมิโนกัวนิดีน (สารประกอบ 4) ยับยั้งการพัฒนาของอาการชักแบบ kindled ที่เหนี่ยวนำด้วยสารเคมีและอาการชักแบบเกิดขึ้นเองซ้ำในหนูเมาส์ในลักษณะขึ้นกับขนาดยา นอกจากนี้ สารยับยั้ง iNOS แบบจำเพาะอย่างสารประกอบ 13 ยังลดการพัฒนาของโรคลมชักกลีบขมับในหนูแรทที่ได้รับสารพิษต่อระบบประสาทไคเนต ในการศึกษานี้ สัตว์ที่ได้รับการรักษามีภาวะไวเกินของเซลล์ประสาทลดลง (การเกิดคลื่นไฟฟ้าแบบชัก) และแสดงการลดลงของอาการชักแบบเกิดขึ้นเองซ้ำถึงร้อยละ 90 (เทียบกับกลุ่มควบคุม) ในช่วงระยะเวลา 6 เดือนภายหลังได้รับสารไคเนต
โดยรวมแล้ว บทบาทของ iNOS ในสมองอาจซับซ้อนกว่าที่เคยเชื่อกันไว้แต่เดิม การศึกษาล่าสุดชี้ว่า iNOS อาจมีบทบาทบางประการในภาวะซึมเศร้าด้วย Montezuma และคณะรายงานว่าทั้งการลบยีน iNOS และการยับยั้ง iNOS แบบจำเพาะ (โดยใช้สารประกอบ 4 หรือ 13) ช่วยลดระยะเวลาการหยุดนิ่งในการทดสอบว่ายน้ำแบบบังคับในหนูเมาส์ ในลักษณะเดียวกับที่ยาต้านซึมเศร้าทำได้ แต่สารยับยั้ง nNOS แบบจำเพาะ (ที่ขนาดยาเดียวกันเมื่อเทียบกับค่า Ki ของมัน) กลับไม่ได้ผลเช่นนั้น สารประกอบ 13 ยังแสดงความสามารถในการย้อนกลับพฤติกรรมคล้ายซึมเศร้าที่เกิดจากความเครียดในหนูเมาส์ ซึ่งสอดคล้องกับผลการศึกษาก่อนหน้านี้ที่ชี้ว่าความเครียดเรื้อรังอาจนำไปสู่การเพิ่มขึ้นของการอักเสบโดยทั่วไป
การสรุปบทบาทของ iNOS ในมะเร็งอย่างครบถ้วนนั้นอยู่นอกเหนือขอบเขตของบทความทบทวนนี้ แม้สารยับยั้ง iNOS จะแสดงผลหลากหลายทั้งในการป้องกันสารก่อมะเร็งทางเคมีและฤทธิ์ต้านมะเร็งในแบบจำลองสัตว์ทดลองก็ตาม ตัวอย่างเช่น ทั้งสารประกอบ 4 และ 6 ช่วยลดอุบัติการณ์และจำนวนของรอยโรคก่อนเป็นมะเร็งในหนูแรทที่ได้รับสารก่อมะเร็งลำไส้ใหญ่แบบจำเพาะ การให้สารยับยั้ง iNOS ร่วมกับสารยับยั้ง COX-2 อย่างเซเลค็อกซิบยิ่งเสริมศักยภาพในการป้องกันมะเร็งมากขึ้น ซึ่งบ่งชี้ว่า iNOS ที่แสดงออกมากเกินไปอาจกำลังควบคุมการอักเสบผ่านทาง COX-2 นอกจากนี้ยังพบผลดีจากการยับยั้ง iNOS ในหนูเมาส์ที่ปลูกถ่ายเนื้องอกมะเร็งผิวหนังของมนุษย์ สารประกอบ 5 ชะลอการเติบโตของเนื้องอกและแสดงฤทธิ์เสริมกันเมื่อให้ร่วมกับซิสพลาติน ในทำนองเดียวกัน สารประกอบ 5, 22 และไอโซไทโอยูเรียหลายชนิดสามารถป้องกันการเปลี่ยนแปลงของเซลล์หลอดลมของหนูแรทให้กลายเป็นเซลล์มะเร็งได้ ทั้งการลดการแสดงออกของยีน iNOS และการใช้สารประกอบ 1, 2 และ 13 สามารถยับยั้งการเพิ่มจำนวนของเซลล์มะเร็งที่มาจากเต้านม การต่ออายุตัวเองของเซลล์ต้นกำเนิดมะเร็ง และการเคลื่อนย้ายของเซลล์ในหลอดทดลองได้ สารประกอบ 1 และ 2 ยังลดปริมาตรของเนื้องอกและการแพร่กระจายไปยังปอดในแบบจำลองหนูเมาส์ที่ปลูกถ่ายมะเร็งเต้านมชนิดสามลบของมนุษย์ โดยฤทธิ์ของสารเหล่านี้ยังเสริมกันกับยาโดซีแทกเซลอีกด้วย
การยับยั้ง iNOS ยังได้รับการศึกษาสำหรับการรักษาการอักเสบเรื้อรังระดับต่ำที่สัมพันธ์กับเบาหวานชนิดที่ 2 ภาวะดื้ออินซูลินที่สัมพันธ์กับโรคอ้วน และภาวะที่เกี่ยวข้อง Perrault และ Marette รายงานว่าหนูที่ถูกน็อกเอาต์ยีน iNOS แม้จะไม่ได้รับการปกป้องจากโรคอ้วนที่เกิดจากอาหาร แต่ก็ได้รับการปกป้องจากภาวะดื้ออินซูลินที่เกี่ยวข้อง และแสดงความทนต่อกลูโคสที่ดีขึ้นและการดูดซึมกลูโคสในกล้ามเนื้อที่เป็นปกติ Noronha และคณะสังเกตผลลัพธ์ที่คล้ายกันในด้านความทนต่ออินซูลิน แต่หนูอ้วนที่ถูกน็อกเอาต์ยีน iNOS ก็ยังคงมีความดันโลหิตสูงพร้อมกับสารอนุมูลอิสระออกซิเจนของหลอดเลือดที่เพิ่มขึ้น ในทำนองเดียวกัน สารประกอบ 5 ช่วยเพิ่มความไวต่ออินซูลินและลดภาวะน้ำตาลในเลือดสูงขณะอดอาหารในหนูเมาส์อ้วนและเป็นเบาหวาน (ซึ่งมี iNOS แสดงออกมากเกินไปในตับ กล้ามเนื้อ และเนื้อเยื่อไขมัน) โรคอ้วนยังสามารถบั่นทอนการทำงานของระบบน้ำเหลืองได้ (อาจผ่านการสะสมของเซลล์อักเสบรอบหลอดน้ำเหลืองที่ผลิต iNOS) และการรักษาหนูอ้วนด้วยสารประกอบ 13 ส่งผลให้การสะสมของเซลล์อักเสบรอบหลอดน้ำเหลืองลดลงและการทำงานของระบบน้ำเหลืองดีขึ้นเล็กน้อย
สุดท้ายนี้ การยับยั้ง iNOS ยังมีศักยภาพในการรักษาโรคระบบทางเดินหายใจบางชนิด หนูเมาส์ที่ขาด iNOS (แต่ไม่ขาด eNOS) ได้รับการปกป้องจากโรคปอดอุดกั้นเรื้อรังที่เกิดจากควันบุหรี่ การรักษาด้วยสารประกอบ 5 สามารถย้อนกลับความเสียหายของปอดจากควันบุหรี่ที่สะสมมาแปดเดือนได้ ภายหลังการให้ยาเป็นเวลาสามเดือน ตามคำกล่าวของ Nathan สำหรับภาวะนี้และภาวะอื่นๆ iNOS อาจกำลัง “เริ่มสูบบุหรี่” เสียเอง
บทสรุป
iNOS ได้รับการพิสูจน์แล้วว่ามีทั้งผลดีและผลเสียต่อร่างกาย NO ที่ผลิตโดย iNOS มีความจำเป็นต่อการตอบสนองต่อการอักเสบตามปกติ ในขณะที่การควบคุมที่ผิดปกติของ iNOS ก็มีความเกี่ยวข้องกับโรคเฉียบพลันและเรื้อรังหลากหลายชนิด ความก้าวหน้าล่าสุดในการศึกษาโครงสร้างและความเข้าใจใหม่เกี่ยวกับการควบคุมการแสดงออกของ iNOS ได้เปิดทางให้เกิดการออกแบบและพัฒนาสารยับยั้ง iNOS ที่มีความจำเพาะและฤทธิ์แรงสูง แม้สารยับยั้ง iNOS จะแสดงศักยภาพที่น่าพึงพอใจอย่างมากในแบบจำลองสัตว์ทดลองสำหรับภาวะช็อกจากการติดเชื้อ ความเจ็บปวด และภาวะอื่นๆ แต่จนถึงปัจจุบันก็ยังคงล้มเหลวในการทดลองทางคลินิก
ความพยายามต่อไปในสาขานี้ควรมุ่งเน้นไปที่การทำความเข้าใจบทบาทที่ซับซ้อนของ iNOS ในระบบชีวภาพ (ทั้งบทบาทที่เป็นอันตรายและบทบาทปกป้อง) และการปรับปรุงทั้งแบบจำลองสัตว์ทดลองและการทดลองทางคลินิกสำหรับโรคที่เกี่ยวข้องกับการอักเสบ ข้อบ่งชี้ใหม่ที่การยับยั้ง iNOS อาจเป็นประโยชน์ ได้แก่ การรักษามะเร็ง โรคความเสื่อมของระบบประสาท และภาวะซึมเศร้า (รวมถึงภาวะอื่นๆ) ข้อบ่งชี้ใหม่เหล่านี้ได้ฟื้นฟูความสนใจในสาขานี้ขึ้นมาอีกครั้ง และอาจนำไปสู่แนวทางการรักษาใหม่ๆ ในอนาคต
เอกสารอ้างอิง (References)
รายการเอกสารอ้างอิงทั้งหมดด้านล่างนี้คงรูปแบบบรรณานุกรมภาษาอังกฤษไว้ตามต้นฉบับ เพื่อความสะดวกในการสืบค้นแหล่งข้อมูลปฐมภูมิ
- Alderton WK, Cooper CE, Knowles RG. Nitric oxide synthases: structure, function and inhibition. Biochem J. 2001;357:593–615. [PubMed: 11463332]
- Abu-Soud HM, Gachhui R, Raushel FM, et al. The ferrous-dioxy complex of neuronal nitric oxide synthase: divergent effects of l-arginine and tetrahydrobiopterin on its stability. J Biol Chem. 1997;272:17349–17353. [PubMed: 9211873]
- Kone BC, Kuncewicz T, Zhang W, et al. Protein interactions with nitric oxide synthases: controlling the right time, the right place, and the right amount of nitric oxide. Am J Physiol – Renal. 2003;285:F178–F190.
- Sharma JN, Al-Omran A, Parvathy SS. Role of nitric oxide in inflammatory diseases. Inflammopharmacology. 2007;15:252–259. [PubMed: 18236016]
- MacMicking J, Xie Q-w, Nathan CNitric oxide and macrophage function. Annu Rev Immunol. 1997;15:323–350. [PubMed: 9143691]
- Yang Y , Yu T, Lian YJ, et al. Nitric oxide synthase inhibitors: a review of patents from 2011 to the present. Expert Opin Ther Pat 2015; 25: 49–68. [PubMed: 25380586]
- Alderton WK, Angell ADR, Craig C, et al. GW274150 and GW273629 are potent and highly selective inhibitors of inducible nitric oxide synthase in vitro and in vivo. Br J Pharmacol. 2005;145:301–312. [PubMed: 15778742]
- Abu-Soud HM, Stuehr DJ. Nitric oxide synthases reveal a role for calmodulin in controlling electron transfer. Proc Natl Acad Sci USA. 1993;90:10769–10772. [PubMed: 7504282]
- Brenman JE, Chao DS, Gee SH, et al. Interaction of nitric oxide synthase with the postsynaptic density protein PSD-95 and α1-syntrophin mediated by PDZ domains. Cell. 1996;84:757–767. [PubMed: 8625413]
- Förstermann U, Sessa WC. Nitric oxide synthases: regulation and function. Eur Heart J. 2012;33:829–837. [PubMed: 21890489]
- Raman CS, Li H, Martásek P, et al. Crystal structure of constitutive endothelial nitric oxide synthase: a paradigm for pterin function involving a novel metal center. Cell. 1998;95:939–950. [PubMed: 9875848]
- Chreifi G, Li H, McInnes CR, et al. Communication between the zinc and tetrahydrobiopterin binding sites in nitric oxide synthase. Biochemistry. 2014;53:4216–4223. [PubMed: 24819538]
- Venema RC, Sayegh HS, Kent JD, et al. Identification, characterization, and comparison of the calmodulin-binding domains of the endothelial and inducible nitric oxide synthases. J Biol Chem. 1996;271:6435–6440. [PubMed: 8626444]
- Hemmens B, Mayer B. Enzymology of nitric oxide synthases In: Titheradge MA, editor. Nitric oxide protocols. Totowa, NJ: Humana Press; 1998 p. 1–32.
- Clapham DE. Calcium signaling. Cell. 2007;131:1047–1058. [PubMed: 18083096]
- Pan J, Burgher KL, Szczepanik AM, et al. Tyrosine phosphorylation of inducible nitric oxide synthase: implications for potential post-translational regulation. Biochem J. 1996;314:889–894. [PubMed: 8615785]
- Noble MA, Munro AW, Rivers SL, et al. Potentiometric analysis of the flavin cofactors of neuronal nitric oxide synthase. Biochemistry. 1999;38:16413–16418. [PubMed: 10600101]
- Campbell MG, Smith BC, Potter CS, et al. Molecular architecture of mammalian nitric oxide synthases. Proc Natl Acad Sci USA. 2014;111:E3614–E3623. [PubMed: 25125509]
- Li W, Chen L, Lu C, et al. Regulatory role of Glu546 in flavin mononucleotide — heme electron transfer in human inducible nitric oxide synthase. Inorg Chem. 2013;52:4795–4801. [PubMed: 23570607]
- Giroud C, Moreau M, Mattioli TA, et al. Role of arginine guanidinium moiety in nitric oxide synthase mechanism of oxygen activation. J Biol Chem. 2010;285:7233–7245. [PubMed: 19951943]
- Mayer B, Wu C, Gorren ACF, et al. Tetrahydrobiopterin binding to macrophage inducible nitric oxide synthase: heme spin shift and dimer stabilization by the potent pterin antagonist 4-amino- tetrahydrobiopterin. Biochemistry. 1997;36:8422–8427. [PubMed: 9204890]
- Crane BR, Arvai AS, Ghosh DK, et al. Structure of nitric oxide synthase oxygenase dimer with pterin and substrate. Science. 1998;279:2121–2126. [PubMed: 9516116]
- Vásquez-Vivar J, Hogg N, Martásek P, et al. Tetrahydrobiopterin-dependent inhibition of superoxide generation from neuronal nitric oxide synthase. J Biol Chem. 1999;274:26736–26742. [PubMed: 10480877]
- Hurshman AR, Krebs C, Edmondson DE, et al. Ability of tetrahydrobiopterin analogues to support catalysis by inducible nitric oxide synthase: formation of a pterin radical is required for enzyme activity. Biochemistry. 2003;42:13287–13303. [PubMed: 14609340]
- Daff S No synthase: Structures and mechanisms. Nitric Oxide. 2010;23:1–11. [PubMed: 20303412]
- Wei C-C, Wang Z-Q, Hemann C, et al. A tetrahydrobiopterin radical forms and then becomes reduced during Nω-hydroxyarginine oxidation by nitric oxide synthase. J Biol Chem. 2003;278:46668–46673. [PubMed: 14504282]
- Wei C-C, Wang Z-Q, Tejero J, et al. Catalytic reduction of a tetrahydrobiopterin radical within nitric oxide synthase. J Bio Chem. 2008;283:11734–11742. [PubMed: 18283102]
- Fischmann TO, Hruza A, Niu XD, et al. Structural characterization of nitric oxide synthase isoforms reveals striking active-site conservation. Nat Struct Mol Biol. 1999;6:233–242.
- Li H, Shimizu H, Flinspach M, et al. The novel binding mode of N-alkyl-N’-hydroxyguanidine to neuronal nitric oxide synthase provides mechanistic insights into no biosynthesis. Biochemistry. 2002;41:13868–13875. [PubMed: 12437343]
- Ghosh S, Wolan D, Adak S, et al. Mutational analysis of the tetrahydrobiopterin-binding site in inducible nitric oxide synthase. J Biol Chem. 1999;274:24100–24112. [PubMed: 10446182]
- Wang Z-Q, Tejero J, Wei C-C, et al. Arg375 tunes tetrahydrobiopterin functions and modulates catalysis by inducible nitric oxide synthase. J Inorg Biochem. 2012;108:203–215. [PubMed: 22173094]
- Naureckiene S, Kodangattil SR, Kaftan EJ, et al. Identification of critical amino acid residues for human iNOS functional activity. Protein J. 2008;27:309–318. [PubMed: 18459037]
- Panda K, Rosenfeld RJ, Ghosh S, et al. Distinct dimer interaction and regulation in nitric oxide synthase types I, II, and III. J Biol Chem. 2002;277:31020–31030. [PubMed: 12048205]
- Li W, Fan W, Chen L, et al. Role of an isoform-specific serine residue in FMN-heme electron transfer in inducible nitric oxide synthase. J Biol Inorg Chem. 2012;17:675–685. [PubMed: 22407542]
- Chakravortty D, Hensel M. Inducible nitric oxide synthase and control of intracellular bacterial pathogens. Microb Infect. 2003;5:621–627.
- Fernhoff NB, Derbyshire ER, Marletta MA. A nitric oxide/cysteine interaction mediates the activation of soluble guanylate cyclase. Proc Natl Acad Sci USA. 2009;106:21602–21607. [PubMed: 20007374]
- Burney S, Caulfield JL, Niles JC, et al. The chemistry of DNA damage from nitric oxide and peroxynitrite. Mutat Res Fund Mol M. 1999;424:37–49.
- Xia C, Misra I, Iyanagi T, et al. Regulation of interdomain interactions by calmodulin in inducible nitric oxide synthase. J Biol Chem. 2009;284:30708–30717. [PubMed: 19737939]
- Denninger JW, Marletta MA. Guanylate cyclase and the NO/cGMP signaling pathway. Biochim Biophys Acta. 1999;1411:334–350. [PubMed: 10320667]
- Chanthaphavong RS, Loughran PA, Lee TYS, et al. A role for cGMP in inducible nitric oxide synthase (iNOS)-induced tumor necrosis factor (TNF) α-converting enzyme (TACE/ADAM17) activation, translocation, and TNF receptor 1 (TNFR1) shedding in hepatocytes. J Biol Chem. 2012;287:35887–35898. [PubMed: 22898814]
- Vannini F, Kashfi K, Nath N. The dual role of iNOS in cancer. Redox Biol. 2015;6:334–343. [PubMed: 26335399]
- Hey C, Boucher J-L, Vadon-Le Goff S, et al. Inhibition of arginase in rat and rabbit alveolar macrophages by Nω-hydroxy-D,L-indospicine, effects on L-arginine utilization by nitric oxide synthase. Br J Pharmacol. 1997;121:395–400. [PubMed: 9179379]
- Ckless K, Lampert A, Reiss J, et al. Inhibition of arginase activity enhances inflammation in mice with allergic airway disease, in association with increases in protein S-nitrosylation and tyrosine nitration. J Immunol. 2008;181:4255–4264. [PubMed: 18768883]
- Bruch-Gerharz D, Schnorr O, Suschek C, et al. Arginase 1 overexpression in psoriasis: limitation of inducible nitric oxide synthase activity as a molecular mechanism for keratinocyte hyperproliferation. Am J Pathol. 2003;162:203–211. [PubMed: 12507903]
- Schmidlin A, Wiesinger H. Transport of L-arginine in cultured glial cells. Glia. 1994;11:262–268. [PubMed: 7960030]
- Bogle RG, Baydoun AR, Pearson JD, et al. L-arginine transport is increased in macrophages generating nitric oxide. Biochem J. 1992;284:15–18. [PubMed: 1599394]
- Manner CK, Nicholson B, MacLeod CL. CAT2 arginine transporter deficiency significantly reduces iNOS-mediated NO production in astrocytes. J Neurochem. 2003;85:476–482. [PubMed: 12675924]
- Kawahara K, Gotoh T, Oyadomari S, et al. Co-induction of argininosuccinate synthetase, cationic amino acid transporter-2, and nitric oxide synthase in activated murine microglial cells. Mol Brain Res. 2001;90:165–173. [PubMed: 11406294]
- Mori M, Gotoh T. Arginine metabolic enzymes, nitric oxide and infection. J Nutr. 2004;134:2820S–2825S. [PubMed: 15465793]
- Cho HJ, Martin E, Xie QW, et al. Inducible nitric oxide synthase: identification of amino acid residues essential for dimerization and binding of tetrahydrobiopterin. Proc Natl Acad Sci USA. 1995;92:11514–11518. [PubMed: 8524794]
- Tzeng E, Billiar TR, Robbins PD, et al. Expression of human inducible nitric oxide synthase in a tetrahydrobiopterin (H4B)-deficient cell line: H4B promotes assembly of enzyme subunits into an active dimer. Proc Natl Acad Sci USA. 1995;92:11771–11775. [PubMed: 8524846]
- Frank S, Pfeilschifter J, Madlener M, et al. Induction of inducible nitric oxide synthase and its corresponding tetrahydrobiopterin-cofactor-synthesizing enzyme GTP-cyclohydrolase I during cutaneous wound repair. J Invest Dermatol. 1998;111:1058–1064. [PubMed: 9856817]
- Youn H, Ji I, Ji HP, et al. Under-expression of kalirin-7 increases iNOS activity in cultured cells and correlates to elevated iNOS activity in Alzheimer’s disease hippocampus. J Alzheimer’s Dis. 2007;12:271–281. [PubMed: 18057561]
- Ratovitski EA, Alam MR, Quick RA, et al. Kalirin inhibition of inducible nitric oxide synthase. J Biol Chem. 1999;274:993–999. [PubMed: 9873042]
- Ratovitski EA, Bao C, Quick RA, et al. An inducible nitric oxide synthase (NOS)-associated protein inhibits NOS dimerization and activity. J Biol Chem. 1999;274:30250–30257. [PubMed: 10514518]
- Zhang ZJ, Cheang LCV , Wang MW, et al. Quercetin exerts a neuroprotective effect through inhibition of the iNOS/NO system and pro-inflammation gene expression in PC12 cells and in zebrafish. Int J Mol Med. 2011;27:195. [PubMed: 21132259]
- Kuncewicz T, Balakrishnan P, Snuggs MB, et al. Specific association of nitric oxide synthase-2 with Rac isoforms in activated murine macrophages. Am J Physiol – Renal. 2001;281:F326–F336.
- Jyoti A, Singh AK, Dubey M, et al. Interaction of inducible nitric oxide synthase with Rac2 regulates reactive oxygen and nitrogen species generation in the human neutrophil phagosomes: Implication in microbial killing. Antioxid Redox Sign. 2013;20:417–431.
- Yang Y , Zhang J, Liu Y , et al. Role of nitric oxide synthase in the development of bone cancer pain and effect of L-NMMA. Mol Med Rep. 2016;13:1220–1226. [PubMed: 26648027]
- Assreuy J, Cunha FQ, Liew FY , et al. Feedback inhibition of nitric oxide synthase activity by nitric oxide. Br J Pharmacol. 1993;108:833–837. [PubMed: 7682140]
- Smith BC, Fernhoff NB, Marletta MA. Mechanism and kinetics of inducible nitric oxide synthase auto-S-nitrosation and inactivation. Biochemistry. 2012;51:1028–1040. [PubMed: 22242685]
- Kleinert H, Schwarz PM, Förstermann U. Regulation of the expression of inducible nitric oxide synthase. Biol Chem. 2003;384:1343–1364. [PubMed: 14669979]
- Pautz A, Art J, Hahn S, et al. Regulation of the expression of inducible nitric oxide synthase. Nitric Oxide. 2010;23:75–93. [PubMed: 20438856]
- Zhou X, Yang W, Li J. Ca2+- and protein kinase C-dependent signaling pathway for nuclear factor- κb activation, inducible nitric oxide synthase expression, and tumor necrosis factor-α production in lipopolysaccharide-stimulated rat peritoneal macrophages. J Biol Chem. 2006;281:31337– 31347. [PubMed: 16923814]
- Titheradge MA. Nitric oxide in septic shock. Biochim Biophys Acta. 1999;1411:437–455. [PubMed: 10320674]
- Ganster RW, Taylor BS, Shao L, et al. Complex regulation of human inducible nitric oxide synthase gene transcription by Stat 1 and NF-κB. Proc Natl Acad Sci USA. 2001;98:8638–8643. [PubMed: 11438703]
- Dell’Albani P, Santangelo R, Torrisi L, et al. Jak/stat signaling pathway mediates cytokine-induced iNOS expression in primary astroglial cell cultures. J Neurosci Res. 2001;65:417–424. [PubMed: 11536325]
- Hevel JM, White KA, Marletta MA. Purification of the inducible murine macrophage nitric oxide synthase. Identification as a flavoprotein. J Biol Chem. 1991;266:22789–22791. [PubMed: 1720773]
- Geller DA, Nussler AK, Di Silvio M, et al. Cytokines, endotoxin, and glucocorticoids regulate the expression of inducible nitric oxide synthase in hepatocytes. Proc Natl Acad Sci USA. 1993;90:522–526. [PubMed: 7678458]
- Nussler AK, Billiar TR, Hoffman RA, et al. Stimulation of the nitric oxide synthase pathway in human hepatocytes by cytokines and endotoxin. J Exp Med. 1992;176:261–264. [PubMed: 1377225]
- Koide M, Kawahara Y , Tsuda T, et al. Cytokine-induced expression of an inducible type of nitric oxide synthase gene in cultured vascular smooth muscle cells. FEBS Lett. 1993;318:213–217. [PubMed: 7680009]
- Macnaul KL, Hutchinson NI. Differential expression of iNOS and cNOS mRNA in human vascular smooth muscle cells and endothelial cells under normal and inflammatory conditions. Biochem Biophys Res Commun. 1993;196:1330–1334. [PubMed: 7504476]
- Charles IG, Palmer RM, Hickery MS, et al. Cloning, characterization, and expression of a cDNA encoding an inducible nitric oxide synthase from the human chondrocyte. Proc Natl Acad Sci USA. 1993;90:11419–11423. [PubMed: 7504305]
- Nomura Y , Kitamura Y . Inducible nitric oxide synthase in glial cells. Neurosci Res. 1993;18:103– [PubMed: 7510374]
- Galea E, Reis DJ, Feinstein DL. Cloning and expression of inducible nitric oxide synthase from rat astrocytes. J Neurosci Res. 1994;37:406–414. [PubMed: 7513765]
- Koprowski H, Zheng YM, Heber-Katz E, et al. In vivo expression of inducible nitric oxide synthase in experimentally induced neurologic diseases. Proc Natl Acad Sci USA. 1993;90:3024– 3027. [PubMed: 7681993]
- Minc-Golomb D, Tsarfaty I, Schwartz JP. Expression of inducible nitric oxide synthase by neurones following exposure to endotoxin and cytokine. Br J Pharmacol. 1994;112:720–722. [PubMed: 7522856]
- Balligand JL, Ungureanu-Longrois D, Simmons WW, et al. Cytokine-inducible nitric oxide synthase (iNOS) expression in cardiac myocytes. Characterization and regulation of iNOS expression and detection of iNOS activity in single cardiac myocytes in vitro. J Biol Chem. 1994;269:27580–27588. [PubMed: 7525557]
- Lyons CR, Orloff GJ, Cunningham JM. Molecular cloning and functional expression of an inducible nitric oxide synthase from a murine macrophage cell line. J Biol Chem. 1992;267:6370– 6374. [PubMed: 1372907]
- Xie Q, Cho H, Calaycay J, et al. Cloning and characterization of inducible nitric oxide synthase from mouse macrophages. Science. 1992;256:225–228. [PubMed: 1373522]
- Albina JE. On the expression of nitric oxide synthase by human macrophages. Why no NO? J Leukocyte Biol. 1995;58:643–649. [PubMed: 7499961]
- Schneemann M, Schoedon G, Hofer S, et al. Nitric oxide synthase is not a constituent of the antimicrobial armature of human mononuclear phagocytes. J Infect Dis. 1993;167:1358–1363. [PubMed: 7684756]
- Geller DA, Lowenstein CJ, Shapiro RA, et al. Molecular cloning and expression of inducible nitric oxide synthase from human hepatocytes. Proc Natl Acad Sci USA. 1993;90:3491–3495. [PubMed: 7682706]
- Obregón E, Punzón MC, González-Nicolás J, et al. Induction of adhesion/differentiation of human neuroblastoma cells by tumour necrosis factor-α requires the expression of an inducible nitric oxide synthase. Eur J Neurosci. 1997;9:1184–1193. [PubMed: 9215702]
- V odovotz YLM, Flanders KC, Chesler L, Xie QW, Smith TW, Weidner J, Mumford R, Webber R, Nathan C, Roberts AB, Lippa CF, and Sporn MB. Inducible nitric oxide synthase in tangle-bearing neurons of patients with Alzheimer’s disease. J Exp Med. 1996;184:1425–1433. [PubMed: 8879214]
- Vannucchi MG, Corsani L, Gianfriddo M, et al. Expression of neuronal and inducible nitric oxide synthase in neuronal and glial cells after transient occlusion of the middle cerebral artery. Neuroscience. 2005;136:1015–1026. [PubMed: 16216429]
- Sparrow JR, Nathan C, V odovotz Y . Cytokine regulation of nitric oxide synthase in mouse retinal pigment epithelial cells in culture. Exp Eye Res. 1994;59:129–139. [PubMed: 7530664]
- Huang H, Rose JL, Hoyt DG. P38 mitogen-activated protein kinase mediates synergistic induction of inducible nitric oxide synthase by lipopolysaccharide and interferon-γ through signal transducer and activator of transcription 1 Ser727 phosphorylation in murine aortic endothelial cells. Mol Pharmacol. 2004;66:302–311. [PubMed: 15266021]
- Williams G, Brown T, Becker L, et al. Cytokine-induced expression of nitric oxide synthase in C2C12 skeletal muscle myocytes. Am J Physiol-Reg I. 1994;267:R1020–R1025.
- Saha RN, Pahan K. Regulation of inducible nitric oxide synthase gene in glial cells. Antioxid Redox Sign. 2006;8:929–947.
- Hu S, Ali H, Sheng WS, et al. Gp-41-mediated astrocyte inducible nitric oxide synthase mrna expression: Involvement of interleukin-1β production by microglia. J Neurosci. 1999;19:6468– 6474. [PubMed: 10414975]
- Liu X, Jana M, Dasgupta S, et al. Human immunodeficiency virus type 1 (HIV-1) tat induces nitric oxide synthase in human astroglia. J Biol Chem. 2002;277:39312–39319. [PubMed: 12167619]
- Majano PL, García-Monzón C, López-Cabrera M, et al. Inducible nitric oxide synthase expression in chronic viral hepatitis. Evidence for a virus-induced gene upregulation. J Clin Invest. 1998;101:1343–1352. [PubMed: 9525976]
- Ochoa JB, Udekwu AO, Billiar TR, et al. Nitrogen oxide levels in patients after trauma and during sepsis. Ann Surg. 1991;214:621–626. [PubMed: 1953116]
- Annane D, Sanquer S, Sébille V , et al. Compartmentalised inducible nitric oxide synthase activity in septic shock. Lancet. 2000;355:1143–1148. [PubMed: 10791377]
- Kobayashi Y , Ikeda K, Shinozuka K, et al. L-nitroarginine increases blood pressure in the rat. Clin Exp Pharmacol Physiol. 1991;18:397–399. [PubMed: 1914242]
- Adamson DC, McArthur JC, Dawson TM, et al. Rate and severity of HIV-associated dementia (HAD): correlations with Gp41 and iNOS. Mol Med. 1999;5:98–109. [PubMed: 10203575]
- Persichini T, Mancino G, Cappelli G, et al. Mycobacterium tuberculosis enhances iNOS mRNA expression and HIV replication in human astrocytoma cells. Neuroreport. 1997;8:1897–1901. [PubMed: 9223073]
- Cramer JP, Mockenhaupt FP, Ehrhardt S, et al. iNOS promoter variants and severe malaria in Ghanaian children. Trop Med Int Health. 2004;9:1074–1080. [PubMed: 15482399]
- Noronha BT, Li J-M, Wheatcroft SB, et al. Inducible nitric oxide synthase has divergent effects on vascular and metabolic function in obesity. Diabetes. 2005;54:1082–1089. [PubMed: 15793247]
- Soskić SS, Dobutović BD, Sudar EM, et al. Regulation of inducible nitric oxide synthase (iNOS) and its potential role in insulin resistance, diabetes and heart failure. Open Card Med J. 2011;5:153–163.
- Kröncke K-D, Brenner H-H, Rodriguez M-L, et al. Pancreatic islet cells are highly susceptible towards the cytotoxic effects of chemically generated nitric oxide. BBA – Molecular Basis Dis. 1993;1182:221–229.
- Kleemann R, Rothe H, Kolb-Bachofen V , et al. Transcription and translation of inducible nitric oxide synthase in the pancreas of prediabetic BB rats. FEBS Lett. 1993;328:9–12. [PubMed: 7688327]
- Carvalho-Filho MA, Ueno M, Carvalheira JBC, et al. Targeted disruption of iNOS prevents LPS- induced S-nitrosation of IR beta/IRS-1 and Akt and insulin resistance in muscle of mice. Am J Physiol – Endoc M. 2006;291:E476–E482.
- Pilon G, Charbonneau A, White PJ, et al. Endotoxin mediated-iNOS induction causes insulin resistance via ONOO− induced tyrosine nitration of IRS-1 in skeletal muscle. PLOS ONE. 2011;5:e15912.
- Granados-Principal S, Liu Y , Guevara ML, et al. Inhibition of iNOS as a novel effective targeted therapy against triple-negative breast cancer. Breast Cancer Res. 2015;17:25. [PubMed: 25849745]
- Lejeune P, Lagadec P, Onier N, et al. Nitric oxide involvement in tumor-induced immunosuppression. J Immunol. 1994;152:5077–5083. [PubMed: 7513729]
- Meller ST, Dykstra C, Grzybycki D, et al. The possible role of glia in nociceptive processing and hyperalgesia in the spinal cord of the rat. Neuropharmacology. 1994;33:1471–1478. [PubMed: 7532831]
- Levy D, Höke A, Zochodne DW. Local expression of inducible nitric oxide synthase in an animal model of neuropathic pain. Neurosci Lett. 1999;260:207–209. [PubMed: 10076904]
- Yoshida M, Xia Y . Heat shock protein 90 as an endogenous protein enhancer of inducible nitric oxide synthase. J Biol Chem. 2003;278:36953–36958. [PubMed: 12855682]
- Kawase M, Kinouchi H, Kato I, et al. Inducible nitric oxide synthase following hypoxia in rat cultured glial cells. Brain Res. 1996;738:319–322. [PubMed: 8955528]
- Haas J, Storch-Hagenlocher B, Biessmann A, et al. Inducible nitric oxide synthase and argininosuccinate synthetase: co-induction in brain tissue of patients with Alzheimer’s dementia and following stimulation with β-amyloid 1–42 in vitro. Neurosci Lett. 2002;322:121–125. [PubMed: 11958858]
- Koppula S, Kumar H, Kim IS, et al. Reactive oxygen species and inhibitors of inflammatory enzymes, NADPH oxidase, and iNOS in experimental models of Parkinson’s disease. Mediat Inflamm. 2012;2012:16.
- Wilcock DM, Lewis MR, Van Nostrand WE, et al. Progression of amyloid pathology to Alzheimer’s disease pathology in an amyloid precursor protein transgenic mouse model by removal of nitric oxide synthase 2. J Neurosci. 2008;28:1537–1545. [PubMed: 18272675]
- Hevel JM, Marletta MA. Nitric oxide synthase assays. Methods Enzymol. 1994;233:250–258. [PubMed: 7516999]
- Naureckiene S, Edris W, Ajit SK, et al. Use of a murine cell line for identification of human nitric oxide synthase inhibitors. J Pharmacol Toxicol Methods. 2007;55:303–313. [PubMed: 16990017]
- Nemzek JA, Hugunin KMS, Opp MR. Modeling sepsis in the laboratory: merging sound science with animal well-being. Comparative Med. 2008;58:120–128.
- Wang LX, Wang ZJ. Animal and cellular models of chronic pain. Adv Drug Del Rev. 2003;55:949–965.
- Winter C, Risley EA, and Nuss GW Carrageenin-induced edema in hind paw of the rat as an assay for antiinflammatory drugs. Proc Soc Exp Biol Med. 1962;111:544–547. [PubMed: 14001233]
- Kim S, and Chung JM An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. PAIN. 1992;50:355–363. [PubMed: 1333581]
- Bennett GJ, Xie YK. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain. 1988;33:87–107. [PubMed: 2837713]
- Whiteley PE, Dalrymple SA. Models of inflammation: adjuvant-induced arthritis in the rat. Current protocols in pharmacology: John Wiley & Sons, Inc; 2001.
- Vítacek J, Lojek, A, Valacchi, G, and Kubala, L. Arginine-based inhibitors of nitric oxide synthase: therapeutic potential and challenges. Mediat Inflamm. 2012;2012:22.
- Olken NM, Osawa Y , Marletta MA. Characterization of the inactivation of nitric oxide synthase by NG-methyl-L-arginine: evidence for heme loss. Biochemistry. 1994;33:14784–14791. [PubMed: 7527657]
- Haynes WG, Noon JP, Walker BR, et al. Inhibition of nitric oxide synthesis increases blood pressure in healthy humans. J Hypertens. 1993;11:1375–1380. [PubMed: 7510736]
- Wolff DJ, Lubeskie A. Aminoguanidine is an isoform-selective, mechanism-based inactivator of nitric oxide synthase. Arch Biochem Biophys. 1995;316:290–301. [PubMed: 7530937]
- Fast W, Nikolic D, Van Breemen RB, et al. Mechanistic studies of the inactivation of inducible nitric oxide synthase by N5-iminoethyl-L-ornithine. J Am Chem Soc. 1999;121:903–916.
- Moore WM, Webber RK, Jerome GM, et al. L-N6-(1-Iminoethyl)lysine: A selective inhibitor of inducible nitric oxide synthase. J Med Chem. 1994;37:3886–3888. [PubMed: 7525961]
- László F, Whittle BJR. Actions of isoform-selective and non-selective nitric oxide synthase inhibitors on endotoxin-induced vascular leakage in rat colon. Eur J Pharmacol. 1997;334:99– [PubMed: 9346334]
- Tang W, Li H, Poulos TL, et al. Mechanistic studies of inactivation of inducible nitric oxide synthase by amidines. Biochemistry. 2015;54:2530–2538. [PubMed: 25811913]
- Corbett JA, Tilton RG, Chang K, et al. Aminoguanidine, a novel inhibitor of nitric oxide formation, prevents diabetic vascular dysfunction. Diabetes. 1992;41:552–556. [PubMed: 1376704]
- Misko TP, Moore WM, Kasten TP, et al. Selective inhibition of the inducible nitric oxide synthase by aminoguanidine. Eur J Pharmacol. 1993;233:119–125. [PubMed: 7682510]
- Tinker A, and Wallace A V Selective inhibitors of inducible nitric oxide synthase: potential agents for the treatment of inflammatory diseases? Curr Top Med Chem. 2006;6:77–92. [PubMed: 16454760]
- Wu CC, Chen SJ, Szabó C, et al. Aminoguanidine attenuates the delayed circulatory failure and improves survival in rodent models of endotoxic shock. Br J Pharmacol. 1995;114:1666–1672. [PubMed: 7541282]
- Cross AH, Misko TP, Lin RF, et al. Aminoguanidine, an inhibitor of inducible nitric oxide synthase, ameliorates experimental autoimmune encephalomyelitis in SJL mice. J Clin Invest. 1994;93:2684–2690. [PubMed: 7515395]
- Wolff DJ, Lubeskie A, Gauld DS, et al. Inactivation of nitric oxide synthases and cellular nitric oxide formation by N6-iminoethyl-L-lysine and N5-iminoethyl-L-ornithine. Eur J Pharmacol. 1998;350:325–334. [PubMed: 9696424]
- Pelletier J-P, Jovanovic DV , Lascau-Coman V , et al. Selective inhibition of inducible nitric oxide synthase reduces progression of experimental osteoarthritis in vivo: Possible link with the reduction in chondrocyte apoptosis and caspase 3 level. Arthritis Rheum. 2000;43:1290–1299. [PubMed: 10857787]
- Hallinan EA, Tsymbalov S, Dorn CR, et al. Synthesis and biological characterization of L-N6-(1- iminoethyl)lysine 5-tetrazole-amide, a produg of a selective iNOS inhibitor. J Med Chem. 2002;45:1686–1689. [PubMed: 11931623]
- Hansel TT, Kharitonov SA, Donnelly LE, et al. A selective inhibitor of inducible nitric oxide synthase inhibits exhaled breath nitric oxide in healthy volunteers and asthmatics. FASEB J. 2003.
- De Alba J, Clayton NM, Collins SD, et al. GW274150, a novel and highly selective inhibitor of the inducible isoform of nitric oxide synthase (iNOS), shows analgesic effects in rat models of inflammatory and neuropathic pain. Pain. 2006;120:170–181. [PubMed: 16360270]
- Dugo L, Marzocco S, Mazzon E, et al. Effects of GW274150, a novel and selective inhibitor of iNOS activity, in acute lung inflammation. Br J Pharmacol. 2004;141:979–987. [PubMed: 14769784]
- Seymour M, Petavy F, Chiesa F, et al. Ultrasonographic measures of synovitis in an early phase clinical trial: a double-blind, randomized, placebo and comparator controlled phase IIa trial of GV274150 (a selective inducible nitric oxide synthase inhibitor) in rheumatoid arthritis. Clin Exp Rheumatol 2012;30:254–261. [PubMed: 22409880]
- Høivik HO, Laurijssens BE, Harnisch LO, et al. Lack of efficacy of the selective iNOS inhibitor GW274150 in prophylaxis of migraine headache. Cephalalgia. 2010;30:1458–1467. [PubMed: 20974604]
- Dunstan RRKhan RWNK Skin wound healing in the SKH-1 female mouse following inducible nitric oxide synthase inhibition. Br J Dermatol 2007;157:656–61 [PubMed: 17672879]
- Connor J, Rogers K, Sunyer T, et al. Efficacy of the selective inducible nitric oxide synthase inhibitor SD-6010 in nonclinical inflammatory, neuropathic, and osteoarthritis pain. Osteoarthr Cartilage. 2012;20:S54–S296.
- Hellio leGraverand M-P, Clemmer RS, Redifer P, et al. A 2-year randomised, double-blind, placebo-controlled, multicentre study of oral selective iNOS inhibitor, cindunistat (SD-6010), in patients with symptomatic osteoarthritis of the knee. Ann Rheum Dis. 2013;72:187–195. [PubMed: 23144445]
- Southan GJ, Szabó C, Thiemermann C. Isothioureas: potent inhibitors of nitric oxide synthases with variable isoform selectivity. Br J Pharmacol. 1995;114:510–516. [PubMed: 7533622]
- Chayah M, Camacho ME, Carrión MD, et al. N,N-disubstituted thiourea and urea derivatives: design, synthesis, docking studies, and biological evaluation against nitric oxide synthase. MedChemComm. 2016;7:667–678.
- Chayah M, Carrión MD, Gallo MA, et al. Development of urea and thiourea kynurenamine derivatives: Synthesis, molecular modeling, and biological evaluation as nitric oxide synthase inhibitors. ChemMedChem. 2015;10:874–882. [PubMed: 25801086]
- Garvey EP, Oplinger JA, Tanoury GJ, et al. Potent and selective inhibition of human nitric oxide synthases. Inhibition by non-amino acid isothioureas. J Biol Chem. 1994;269:26669–26676. [PubMed: 7523409]
- Garvey EP, Oplinger JA, Furfine ES, et al. 1400W is a slow, tight binding, and highly selective inhibitor of inducible nitric oxide synthase in vitro and in vivo. J Biol Chem. 1997;272:4959– 4963. [PubMed: 9030556]
- Zhu Y , Nikolic D, Van Breemen RB, et al. Mechanism of inactivation of inducible nitric oxide synthase by amidines. Irreversible enzyme inactivation without inactivator modification. J Am Chem Soc. 2005;127:858–868. [PubMed: 15656623]
- Staunton C, Djouhri L, Barrett-Jolley R, and Thippeswami T 1400W alleviates pain and increases the expression levels of anti-nociceptive and neuromodulatory mediators. P Physiol Soc. 2014:31: p126P, PCA151, poster communications.
- Maccallini C, Patruno A, Bešker N, et al. Synthesis, biological evaluation, and docking studies of N-substituted acetamidines as selective inhibitors of inducible nitric oxide synthase. J Med Chem. 2009;52:1481–1485. [PubMed: 19203270]
- Moore WM, Webber RK, Fok KF, et al. 2-iminopiperidine and other 2-iminoazaheterocycles as potent inhibitors of human nitric oxide synthase isoforms. J Med Chem. 1996;39:669–672. [PubMed: 8576908]
- Naka M, Nanbu T, Kobayashi K, et al. A potent inhibitor of inducible nitric oxide synthase, ONO-1714, a cyclic amidine derivative. Biochem Biophys Res Commun. 2000;270:663–667. [PubMed: 10753680]
- Segikuchi F, Mita Y , Kamanaka Y , Kawao N, Matsuya H, Taga C, and Kawabata A The potent inducible nitric oxide synthase inhibitor ONO-1714 inhibits neuronal NOS and exerts antinociception in rats. Neurosci. Lett. 365 (2), 111–115.
- Mikawa K, Nishina K, Takao Y , et al. ONO-1714, a nitric oxide synthase inhibitor, attenuates endotoxin-induced acute lung injury in rabbits. Anesth Analg. 2003;97:1751–1755. [PubMed: 14633554]
- Mitaka C, Hirata Y , Yokoyama K, et al. A selective inhibitor for inducible nitric oxide synthase improves hypotension and lactic acidosis in canine endotoxic shock. Crit Care Med. 2001;29:2156–2161. [PubMed: 11700413]
- Hagen TJ, Bergmanis AA, Kramer SW, et al. 2-Iminopyrrolidines as potent and selective inhibitors of human inducible nitric oxide synthase. J Med Chem. 1998;41:3675–3683. [PubMed: 9733492]
- Tsymbalov S, Hagen TJ, Moore WM, et al. 3-Hydroxy-4-methyl-5-pentyl-2-iminopyrrolidine: A potent and highly selective inducible nitric oxide synthase inhibitor. Bioorg Med Chem Lett. 2002;12:3337–3339. [PubMed: 12392746]
- Beaton H, Hamley P, Nicholls DJ, et al. 3,4-Dihydro-1-isoquinolinamines: a novel class of nitric oxide synthase inhibitors with a range of isoform selectivity and potency. Bioorg Med Chem Lett. 2001;11:1023–1026. [PubMed: 11327580]
- Tinker AC, Beaton HG, Boughton-Smith N, et al. 1,2-Dihydro-4-quinazolinamines: potent, highly selective inhibitors of inducible nitric oxide synthase which show antiinflammatory activity in vivo. J Med Chem. 2003;46:913–916. [PubMed: 12620067]
- LaBuda CJ, Koblish M, Tuthill P, et al. Antinociceptive activity of the selective iNOS inhibitor AR-C102222 in rodent models of inflammatory, neuropathic and post-operative pain. Eur J Pain. 2006;10:505–505. [PubMed: 16125426]
- Crane BR, Arvai AS, Gachhui R, et al. The structure of nitric oxide synthase oxygenase domain and inhibitor complexes. Science. 1997;278:425–431. [PubMed: 9334294]
- Garcin ED, Arvai AS, Rosenfeld RJ, et al. Anchored plasticity opens doors for selective inhibitor design in nitric oxide synthase. Nat Chem Biol. 2008;4:700–707. [PubMed: 18849972]
- Hagmann WK, Caldwell CG, Chen P, et al. Substituted 2-aminopyridines as inhibitors of nitric oxide synthases. Bioorg Med Chem Lett. 2000;10:1975–1978. [PubMed: 10987430]
- Pettipher E, Hibbs T, Smith M, et al. Analgesic activity of 2-amino-4-methylpyridine, a novel NO synthase inhibitor. Inflammation Res. 1997;46:135–136.
- Connolly S, Aberg A, Arvai A, et al. 2-aminopyridines as highly selective inducible nitric oxide synthase inhibitors. Differential binding modes dependent on nitrogen substitution. J Med Chem. 2004;47:3320–3323. [PubMed: 15163211]
- Strub A, Ulrich W-R, Hesslinger C, et al. The novel 2-[2-(4-methoxy-pyridin-2-yl)-ethyl]-3H- imidazo[4,5-b]pyridine (BYK191023) is a highly selective inhibitor of the inducible nitric oxide synthase. Mol Pharmacol. 2006;69:328–337. [PubMed: 16223957]
- Tiso M, Strub A, Hesslinger C, et al. BYK191023 (2-[2-(4-methoxy-pyridin-2-yl)-ethyl]- 3H- imidazo[4,5-b]pyridine is an NADPH- and time-dependent irreversible inhibitor of inducible nitric oxide synthase. Mol Pharmacol. 2008;73:1244–1253. [PubMed: 18178668]
- Lehner MD, Marx D, Boer R, et al. In vivo characterization of the novel imidazopyridine BYK191023 (2-[2-(4-methoxy-pyridin-2-yl)-ethyl]-3H-imidazo[4,5-b]pyridine), a potent and highly selective inhibitor of inducible nitric oxide synthase. J Pharmacol Exp Ther. 2006;317:181–187. [PubMed: 16368897]
- Su F, Huang H, Akieda K, et al. Effects of a selective iNOS inhibitor versus norepinephrine in the treatment of septic shock. Shock. 2010;34:243–249. [PubMed: 20160666]
- Sennequier N, Wolan D, Stuehr DJ. Antifungal imidazoles block assembly of inducible NO synthase into an active dimer. J Biol Chem. 1999;274:930–938. [PubMed: 9873034]
- McMillan K, Adler M, Auld DS, et al. Allosteric inhibitors of inducible nitric oxide synthase dimerization discovered via combinatorial chemistry. Proc Natl Acad Sci USA. 2000;97:1506– 1511. [PubMed: 10677491]
- Blasko E, Glaser CB, Devlin JJ, et al. Mechanistic studies with potent and selective inducible nitric oxide synthase dimerization inhibitors. J Biol Chem. 2002;277:295–302. [PubMed: 11689556]
- Nagpal L, Haque MM, Saha A, et al. Mechanism of inducible nitric oxide synthase dimerization inhibition by novel pyrimidine imidazoles. J Biol Chem. 2013;288:19685–19697. [PubMed: 23696643]
- Davey DD, Adler M, Arnaiz D, et al. Design, synthesis, and activity of 2-imidazol-1-ylpyrimidine derived inducible nitric oxide synthase dimerization inhibitors. J Med Chem. 2007;50:1146– 1157. [PubMed: 17315988]
- Bonnefous C, Payne JE, Roppe J, et al. Discovery of inducible nitric oxide synthase (iNOS) inhibitor development candidate KD7332, part 1: identification of a novel, potent, and selective series of quinolinone iNOS dimerization inhibitors that are orally active in rodent pain models. J Med Chem. 2009;52:3047–3062. [PubMed: 19374401]
- Symons KT, Massari ME, Nguyen PM, et al. KLYO956 is a non-imidazole-based orally active inhibitor of nitric oxide synthase dimerization. Mol Pharmacol. 2009;76:153–162. [PubMed: 19364813]
- Payne JE, Bonnefous C, Symons KT, et al. Discovery of dual inducible/neuronal nitric oxide synthase (iNOS/nNOS) inhibitor development candidate 4-((2-cyclobutyl-1h-imidazo[4,5- b]pyrazin-1-yl)methyl)-7,8-difluoroquinolin-2(1h)-one (kd7332) part 2: Identification of a novel, potent, and selective series of benzimidazole-quinolinone iNOS/nNOS dimerization inhibitors that are orally active in pain models. J Med Chem. 2010;53:7739–7755. [PubMed: 20931971]
- Kalypsys I iNOS inhibitor, topical pain product, KD7040, http://www.kalypsys.com/pipeline/ clinical/kd7040.pdf. accessed April 12th, 2017, also available at: https://web.archive.org/web/ 20070327050804/http://www.kalypsys.com/pipeline/clinical/kd7040.pdf).
- Cowans B, Mougin-Andres P, Wheeler P, et al. Salts of inducible nitric oxide synthase dimerization inhibitors. Pat Appl US20070197609. 8, 2007.
- Lopez A, Lorente JA, Steingrub J, et al. Multiple-center, randomized, placebo-controlled, double- blind study of the nitric oxide synthase inhibitor 546C88: Effect on survival in patients with septic shock. Crit Care Med 2004;32:21–30. [PubMed: 14707556]
- Yamashita T, Kawashima S, Ohashi Y , et al. Resistance to endotoxin shock in transgenic mice overexpressing endothelial nitric oxide synthase. Circulation. 2000;101:931–937. [PubMed: 10694534]
- Stahl W, Matejovic M, Radermacher P. Inhibition of nitric oxide synthase during sepsis: revival because of isoform selectivity? Shock. 2010;34:321–322. [PubMed: 20601934]
- Cauwels A, Van Molle W, Janssen B, et al. Protection against TNF-induced lethal shock by soluble guanylate cyclase inhibition requires functional inducible nitric oxide synthase. Immunity. 2000;13:223–231. [PubMed: 10981965]
- Bredan AS, Cauwels A. Is there no treatment for severe sepsis? Libyan J Med. 2008;3:34–38. [PubMed: 21499479]
- Mogil JS. Animal models of pain: progress and challenges. Nat Rev Neurosci. 2009;10:283–294. [PubMed: 19259101]
- Mungrue IN, Gros R, You X, et al. Cardiomyocyte overexpression of iNOS in mice results in peroxynitrite generation, heart block, and sudden death. J Clin Inv. 2002;109:735–743.
- Alexander J, Reynolds HR, Stebbins AL, et al. Effect of tilarginine acetate in patients with acute myocardial infarction and cardiogenic shock: The TRIUMPH randomized controlled trial. JAMA. 2007;297:1657–1666. [PubMed: 17387132]
- Bailey A, Pope TW, Moore SA, et al. The tragedy of TRIUMPH for nitric oxide synthesis inhibition in cardiogenic shock. Am J Cardiovasc Drugs. 2007;7:337–345. [PubMed: 17953472]
- Howes LG, Brillante DG. Expert opinion on tilarginine in the treatment of shock. Expert Opin Inv Drugs. 2008;17:1573–1580.
- Dokken BB, Gaballa MA, Hilwig RW, et al. Inhibition of nitric oxide synthases, but not inducible nitric oxide synthase, selectively worsens left ventricular function after successful resuscitation from cardiac arrest in swine. Acad Emerg Med. 2015;22:197–203. [PubMed: 25639298]
- Mukherjee P, Cinelli MA, Kang S, et al. Development of nitric oxide synthase inhibitors for neurodegeneration and neuropathic pain. Chem Soc Rev. 2014;43:6814–6838. [PubMed: 24549364]
- Li M, Dai F-r, Du X-p, et al. Neuroprotection by silencing iNOS expression in a 6-OHDA model of Parkinson’s disease. J Mol Neurosci. 2012;48:225–233. [PubMed: 22638860]
- Tsuji M, Higuchi Y , Shiraishi K, et al. Protective effect of aminoguanidine on hypoxic-ischemic brain damage and temporal profile of brain nitric oxide in neonatal rat. Pediatr Res. 2000;47:79– [PubMed: 10625086]
- Cockroft KM, Meistrell M, Zimmerman GA, et al. Cerebroprotective effects of aminoguanidine in a rodent model of stroke. Stroke. 1996;27:1393–1398. [PubMed: 8711808]
- Broom L, Marinova-Mutafchieva L, Sadeghian M, et al. Neuroprotection by the selective iNOS inhibitor GW274150 in a model of Parkinson disease. Free Radical Biol Med. 2011;50:633–640. [PubMed: 21185368]
- Zhong H-J, Liu L-J, Chong C-M, et al. Discovery of a natural product-like iNOS inhibitor by molecular docking with potential neuroprotective effects in vivo. PLOS ONE. 2014;9:e92905. [PubMed: 24690920]
- Üzüm G, Akgün-Dar K, Kandil A, et al. Similarity of inflammatory response in epileptic seizures and sepsis: Does the sensitivity to sepsis in epileptic patients increase? Crit Care. 2014;18:P56– P56.
- Rehni AK, Singh TG, Kalra R, et al. Pharmacological inhibition of inducible nitric oxide synthase attenuates the development of seizures in mice. Nitric Oxide. 2009;21:120–125. [PubMed: 19559095]
- Puttachary S, Sharma S, Verma S, et al. 1400W, a highly selective inducible nitric oxide synthase inhibitor is a potential disease modifier in the rat kainate model of temporal lobe epilepsy. Neurobiol Dis. 2016;93:184–200. [PubMed: 27208748]
- Montezuma K, Biojone C, Lisboa SF, et al. Inhibition of iNOS induces antidepressant-like effects in mice: pharmacological and genetic evidence. Neuropharmacology. 2012;62:485–491. [PubMed: 21939674]
- Peng Y-L, Liu Y-N, Liu L, et al. Inducible nitric oxide synthase is involved in the modulation of depressive behaviors induced by unpredictable chronic mild stress. J Neuroinflamm. 2012;9:75– 75.
- Janakiram NB, Rao CV . iNOS-selective inhibitors for cancer prevention: promise and progress. Future Med Chem. 2012;4:2193–2204. [PubMed: 23190107]
- Rao CV , Indranie C, Simi B, et al. Chemopreventive properties of a selective inducible nitric oxide synthase inhibitor in colon carcinogenesis, administered alone or in combination with celecoxib, a selective cyclooxygenase-2 inhibitor. Cancer Res. 2002;62:165–170. [PubMed: 11782374]
- Sikora AG, Gelbard A, Davies MA, et al. Targeted inhibition of inducible nitric oxide synthase inhibits growth of human melanoma in vivo and synergizes with chemotherapy. Clin Cancer Res. 2010;16:1834–1844. [PubMed: 20215556]
- Sharma S, Wilkinson BP, Gao P, et al. Differential activity of NO synthase inhibitors as chemopreventive agents in a primary rat tracheal epithelial cell transformation system. Neoplasia (New York, NY). 2002;4:332–336.
- Perreault M, Marette A. Targeted disruption of inducible nitric oxide synthase protects against obesity-linked insulin resistance in muscle. Nat Med. 2001;7:1138–1143. [PubMed: 11590438]
- Fujimoto M, Shimizu N, Kunii K, et al. A role for iNOS in fasting hyperglycemia and impaired insulin signaling in the liver of obese diabetic mice. Diabetes. 2005;54:1340–1348. [PubMed: 15855318]
- Torrisi JS, Hespe GE, Cuzzone DA, et al. Inhibition of inflammation and iNOS improves lymphatic function in obesity. Sci Rep. 2016;6:19817. [PubMed: 26796537]
- Seimetz M, Parajuli N, Pichl A, et al. Inducible NOS inhibition reverses tobacco-smoke-induced emphysema and pulmonary hypertension in mice. Cell. 2011;147:293–305. [PubMed: 22000010]
- Nathan C Is iNOS beginning to smoke? Cell 147:257–258. [PubMed: 22000003]